Œ‹‰ت‚¨‚و‚رچlژ@
“yڈëƒVپ[ƒhƒoƒ“ƒN‚ج‰ًگح
ƒˆƒ‚ƒM‘گŒ´‚ج“yڈë‚©‚ç19ژي(ٹO—ˆ4ژيپAچف—ˆ15ژي)پA—ر“à‚ج“yڈë‚©‚ç21ژي(ٹO—ˆ6ژيپAچف—ˆ15ژي)پAگى•س‚ج“yڈë‚©‚ç5ژي(ٹO—ˆ2ژيپAچف—ˆ3ژي)‚ج”‰è‚ھ‚ف‚ç‚êپA‘S‘ج‚إ30ژيپiٹO—ˆ9ژيپAچف—ˆ21ژيپj‚ج”‰è‚ھٹm”F‚³‚ꂽپBƒˆƒ‚ƒM‘گŒ´‚ج“yڈë‚ة‚¨‚¢‚ؤ”‰èŒآ‘جگ”‚ھ‘½‚©‚ء‚½ژي‚حپAƒVƒچƒcƒپƒNƒTپi115Œآ‘جپjپAƒ`ƒKƒ„پi85پjپAƒJƒ^ƒoƒ~پi38پjپAƒJƒ‚ƒKƒ„پi26پj‚إ‚ ‚ء‚½پiƒVƒچƒcƒپƒNƒTپAƒJƒ‚ƒKƒ„‚حٹO—ˆگA•¨پjپB—ر“à‚ج“yڈë‚ة‚¨‚¢‚ؤ”‰èŒآ‘جگ”‚ھ‘½‚©‚ء‚½ژي‚حپAƒ„ƒGƒ€ƒOƒ‰پi64Œآ‘جپjپAƒnƒRƒxپi52پj‚إ‚ ‚ء‚½پBگى•س‚ج“yڈë‚ة‚¨‚¢‚ؤ”‰èŒآ‘جگ”‚ھ‘½‚©‚ء‚½ژي‚حپAƒXƒQsp.پi424Œآ‘جپjپAƒIƒIƒCƒkƒ^ƒfپi239پjپAƒJƒ‚ƒKƒ„پi39‚إ‚ ‚ء‚½پiƒJƒ‚ƒKƒ„‚حٹO—ˆگA•¨پjپB
2003”N“x‚جŒ»’n‚إ‚جگAگ¶’²چ¸پiگ¯–ى2003پj‚ة‚و‚ê‚خپA2003”N“x‚جƒˆƒ‚ƒM‘گŒ´‚ة‚¨‚¯‚éƒVƒچƒcƒپƒNƒT‚ج”ي“xپi–ٌ34.5%پj‚حپAƒˆƒ‚ƒM‚ج”ي“xپi–ٌ4.7%پj‚ج–ٌ7”{‚إ‚ ‚ء‚½پB–{‰ًگح‚ة‚¨‚¢‚ؤ‚حپAƒˆƒ‚ƒM‘گŒ´‚ج“yڈë‚ة‚¨‚¯‚éƒVƒچƒcƒپƒNƒT‚ج”‰èŒآ‘جگ”پi115پj‚حپAƒˆƒ‚ƒM‚ج”‰èŒآ‘جگ”پi9پj‚ج–ٌ13”{‚إ‚ ‚ء‚½پBƒˆƒ‚ƒM‘گŒ´‚ة‚¨‚¢‚ؤƒˆƒ‚ƒM‚جگ¶ˆç‚ً‘£گi‚·‚邽‚ك‚ة‚حپAٹg‘ه‚µ‚ؤ‚µ‚ـ‚ء‚½ƒVƒچƒcƒپƒNƒT‚جڈœ‹ژ‚ھ•K—v‚إ‚ ‚éپB‚µ‚©‚µپA–{‰ًگح‚ة‚و‚èپAƒVƒچƒcƒپƒNƒT‚ھ‘ه‚«‚بƒVپ[ƒhƒoƒ“ƒN‚ًŒ`گ¬‚µ‚ؤ‚¢‚é‰آ”\گ«‚ھژ¦چ´‚³‚ꂽ‚±‚ئ‚©‚çپAƒVƒچƒcƒپƒNƒTŒآ‘ج‚ًˆّ‚«”²‚¢‚ؤ‚¢‚邾‚¯‚إ‚حپA–{ژي‚ً’Zژٹش‚إƒrƒIƒgپ[ƒv‚©‚çڈœ‹ژ‚·‚邱‚ئ‚ح‚إ‚«‚ب‚¢‚ئچl‚¦‚ç‚ê‚éپB‚µ‚©‚µپAŒp‘±“I‚ةŒآ‘ج‚ًڈœ‹ژ‚·‚ê‚خپAƒVپ[ƒhƒoƒ“ƒN‚حڈk‘ق‚µ‚ؤ‚¢‚‚ئ‚àچl‚¦‚ç‚ê‚éپB
—ر“à“yڈë‚ة‚¨‚¢‚ؤ”‰è‚ھٹm”F‚³‚ꂽƒnƒCƒ`ƒSƒUƒT‚حپAگAگ¶’²چ¸‚إ‚àگ¶ˆç‚ھٹm”F‚³‚ê‚ؤ‚¨‚èپAچ،ŒمƒrƒIƒgپ[ƒv“à‚إ’è’…¥ٹg‘ه‚µ‚ؤ‚¢‚‚à‚ج‚ئٹْ‘ز‚³‚ê‚éپB“¯—l‚ة“yڈëƒVپ[ƒhƒoƒ“ƒNپEگAگ¶’²چ¸‚ج‘o•û‚إٹm”F‚³‚ꂽژي‚حپAƒAƒپƒٹƒJƒtƒEƒچپAƒIƒ‰ƒ“ƒ_ƒ~ƒ~ƒiƒOƒTپAƒnƒRƒxپAƒ„ƒGƒ€ƒOƒ‰‚إ‚ ‚éپB‚±‚ê‚ç‚جژي‚àپAƒrƒIƒgپ[ƒv“à‚ة’è’…¥ٹg‘ه‚µ‚ؤ‚¢‚‰آ”\گ«‚ھچ‚‚¢‚ئچl‚¦‚ç‚ê‚éپBƒAƒپƒٹƒJƒtƒEƒچ‚ئƒIƒ‰ƒ“ƒ_ƒ~ƒ~ƒiƒOƒT‚حٹO—ˆژي‚إ‚ ‚é‚ج‚إپAچ،Œم’چˆسگ[‚“®‘ش‚ً’اگص‚·‚é•K—v‚ھ‚ ‚éپBگى•س‚ةگAچح‚³‚ꂽƒKƒ}‚حپAگى•س‚ج“yڈë‚ة‚¨‚¢‚ؤ‚ح”‰è‚ھٹm”F‚³‚ꂸپAƒˆƒ‚ƒM‘گŒ´‚ئ—ر“à‚ج“yڈë‚ة‚¨‚¢‚ؤ”‰è‚ھٹm”F‚³‚ꂽپBƒKƒ}‚ھ•—ژU•zژيژq‚إ‚ ‚èپA‹كگع‚·‚é’nˆو‚ةژيژq‚ھ”ٍ‚ٌ‚¾‚à‚ج‚ئچl‚¦‚ç‚ê‚éپB
2003”N“x‚ـ‚إ‚ةƒrƒIƒgپ[ƒv“à‚إ‚جگ¶ˆç‚ھٹm”F‚³‚ꂽگA•¨ƒٹƒXƒg‚ئچ،‰ٌ“yڈëƒVپ[ƒhƒoƒ“ƒN‚©‚甉è‚ھٹm”F‚³‚ꂽگA•¨ƒٹƒXƒg‚ً”نٹr‚µ‚½پi•\4~7پjپBچ،”N“x‚ـ‚إ‚جگA•¨‘ٹ’²چ¸‚إگ¶ˆç‚ھˆê“x‚àٹm”F‚³‚ê‚ؤ‚¢‚ب‚¢‚ة‚à‚©‚©‚ي‚炸پA“yڈë‚©‚ç‚ج”‰è‚ھٹm”F‚³‚ꂽژي‚حپAƒGƒmƒLƒOƒTپAƒ€ƒVƒNƒT‚إ‚ ‚ء‚½پB‚±‚ê‚ç‚حƒrƒIƒgپ[ƒvژü•س‚©‚ç•—‚â’¹پA‚à‚µ‚‚حگlˆ×“I‚بˆع“®‚ة‚و‚èژيژq‚ھ“±“ü‚³‚ꂽ‚ئچl‚¦‚ç‚êپA‚±‚ê‚ç‚ح‚·‚ׂؤچف—ˆژي‚إ‚ ‚é‚ج‚إپAچ،ŒمŒآ‘جگ”‚ج‘‰ء‚ئ“yڈëƒVپ[ƒhƒoƒ“ƒN‘‰ء‚ج‰آ”\گ«‚ھٹْ‘ز‚³‚ê‚éپBڈvچH’¼Œم‚ج2001پA2002”N“x‚ةƒrƒIƒgپ[ƒv“à‚إگ¶ˆç‚ھٹm”F‚³‚ꂽ‚ھپA2003”N“x‚ة‚حگ¶ˆç‚ھٹm”F‚³‚ꂸپAچ،‰ٌ“yڈë‚ة‚¨‚¢‚ؤ”‰è‚ھٹm”F‚³‚ꂽژي‚حپAƒVƒچƒUپAƒXƒJƒVƒ^ƒSƒ{ƒE‚إ‚ ‚ء‚½پB‚µ‚©‚µپA“yڈë‚©‚ç‚ج”‰èŒآ‘جگ”‚حڈ‚ب‚¢پiƒVƒچƒU2Œآ‘جپAƒXƒJƒVƒ^ƒSƒ{ƒE6Œآ‘جپj‚ج‚إپAƒVپ[ƒhƒoƒ“ƒN‚حڈ¬‚³‚¢‚ئگ„ژ@‚³‚ê‚éپB
“yڈë‚ة‚¨‚¯‚éٹO—ˆگA•¨‚جڈoŒ»ژيگ”‚ئŒآ‘جگ”‚حپAƒˆƒ‚ƒM‘گŒ´‚إ4ژيچ‡Œv148Œآ‘جپA—ر“à‚إ6ژيچ‡Œv24Œآ‘جپAگى•س‚إ2ژيچ‡Œv41Œآ‘ج‚إ‚ ‚ء‚½پB—ر“à‚ج“yڈë‚ة‚¨‚¢‚ؤ‚حپAڈoŒ»ژيگ”‚ح‘¼‚ج—§’n‚و‚è‚à‘½‚©‚ء‚½‚ھپAŒآ‘جگ”‚ھڈ‚ب‚©‚ء‚½پB‚±‚ê‚حگAچح‚³‚ꂽژ÷–ط‚ھ‘ه‚«‚ˆç‚ء‚ؤ—ر“à‚ً”ي‰A‚µ‚ؤپAٹO—ˆژي‚جگ¶ˆç‚ً—}گ§‚µ‚ؤ‚¢‚邽‚ك‚ئچl‚¦‚ç‚ê‚éپBٹO—ˆگA•¨‚ج‘½‚‚حپAک—گ‚ج‚ ‚é–¾‚é‚¢ڈêڈٹ‚ًچD‚فپA‰èگ¶‚¦‚جژٹْ‚جگ¬’·‚ة–¾‚é‚¢ŒُڈًŒڈ‚ً•K—v‚ئ‚·‚é‚ئچl‚¦‚ç‚ê‚ؤ‚¢‚éپiکh’J¥گX–{1993پjپBژہچغپAچ،”N“x‚ح—ر“à‚جŒُٹآ‹«‚ھٹO—ˆژي‚ة“K‚³‚ب‚¢ˆأ‚¢Œُٹآ‹«پi‘ٹ‘خŒُ—تژq–§“x‚إ10~20%’ِ“xپj‚ئ‚ب‚ء‚ؤ‚¢‚½پi•\14پAگ}51ژQڈئپjپB
”‰è‚ج‰·“xˆث‘¶گ«‰ًگح
پ@ƒAƒhƒoƒ“ƒeƒXƒgƒrƒIƒgپ[ƒv“à‚إچجژو‚µ‚½پA21ژي‚جگA•¨‚جژيژq‚جپAژي•ت‚ج”‰è‚ج‰·“xˆث‘¶گ«‚حپAˆب‰؛‚ج‚ئ‚¨‚è‚ئ‚ب‚ء‚½پB
1.ƒAƒپƒٹƒJƒCƒkƒzƒEƒYƒL(ƒiƒX‰بˆê”Nگ¶‘گ–{پASolanum americanum Mill.)
پ@–{ژي‚ح–kƒAƒپƒٹƒJŒ´ژY‚إپA“ْ–{‚ة‚ح1950”N‘م‚©‚çگN“ü‹Lک^‚ھ‚ ‚éپBŒ»چفپA‘Sچ‘‚ج”¨’n‚âکH–T‚ب‚ا‚ة’è’…‚µ‚ؤ‚¢‚é‚ئ‚³‚ê‚éپiگ´گ…¥گX“c¥œA“c2001پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA‰·“x‚ھچ‚‚‚ب‚é‚ة‚آ‚ê‚ؤچ‚‚‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/6پژ‚إ‚ح‘S‚”‰è‚µ‚ب‚©‚ء‚½‚ج‚ة‘خ‚µ‚ؤپA17/8پژ‚إ‚حچإڈI”‰è—¦‚ھ•½‹د‚إ–ٌ69%‚إ‚ ‚èپA22/10پA25/13پA30/15پژ‚ة‚¨‚¢‚ؤ‚ح100%‚ھ”‰è‚µ‚½(•\8پAگ}4پAگ}27)پB
پ@‚±‚ج‚و‚¤‚ب‰·“xˆث‘¶گ«‚ً—L‚·‚éژيژq‚ًگ¶ژY‚·‚éگA•¨‚حپA’n‰·‚ج’ل‚¢“~‹G‚ة‚ح”‰è‚¹‚¸‚ةپAڈt‚ة‚ب‚ء‚ؤ’n‰·‚ھچ‚‚‚ب‚ء‚ؤ‚©‚甉肵پAگ¶’·‚ًٹJژn‚·‚é‚à‚ج‚ئچl‚¦‚ç‚ê‚éپBژہچغپA–{ƒrƒIƒgپ[ƒv‚إ‚ح‰ؤچ ‚ة‚ب‚ء‚ؤŒآ‘ج‚ھ“÷ٹل‚إٹm”F‚³‚ê‚ؤ‚¢‚éپiگ¯–ى2003پjپB10پژˆب‰؛‚ج’ل‰·‚إ”‰è‚ھٹ®‘S‚ة—}گ§‚³‚ê‚邱‚ئ‚©‚çپA–{ژي‚ح‘پڈt‚ج–¾‚é‚¢—§’nپپ‹Gگك“IƒMƒƒƒbƒv‚ً—ک—p‚إ‚«‚ب‚¢ژي‚ئچl‚¦‚ç‚ê‚éپB‚±‚ج‚½‚ك–{ژي‚حپAک—گ’n‚ب‚ا‚ج—‡’n‚ة‘½‚گ¶ˆç‚µپA—ر“à‚ب‚ا’n‰·‚جڈمڈ¸‚ھ—}گ§‚³‚ê‚é—§’n‚إ‚ح‚ ‚ـ‚èگ¶ˆç‚µ‚ب‚¢‚ئچl‚¦‚ç‚ê‚éپB
2.ƒCƒgƒoƒMƒN(ƒLƒN‰بˆê”Nگ¶‘گ–{پASchkuhria pinnata Kuntze.)
پ@–{ژي‚حƒپƒLƒVƒRŒ´ژY‚إپA“ْ–{‚ة‚ح‘و“ٌژںگ¢ٹE‘هگيŒم‚©‚çگN“ü‹Lک^‚ھ‚ ‚éپiگ´گ…¥گX“c¥œA“c2001پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA25/13پژ‚إچإ‘ه‚ئ‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/6پژ‚إ‚ح•½‹د‚إ62%پA17/8پژ‚إ‚ح–ٌ83%پA22/10‚إ‚ح–ٌ89%پA25/13‚إ‚ح–ٌ97%‚إ‚ ‚èپA30/15پژ‚إ‚ح–ٌ61%‚ھ”‰è‚µ‚½(•\8پAگ}5پAگ}26)پB
پ@‚±‚ج‚و‚¤‚ب‰·“xˆث‘¶گ«‚ً—L‚·‚éژيژq‚ًگ¶ژY‚·‚éگA•¨‚حپA’n‰·‚ج’ل‚¢“~‹G‚ة‚ح”‰è‚¹‚¸‚ةپAڈt‚ة‚ب‚ء‚ؤ’n‰·‚ھچ‚‚‚ب‚ء‚ؤ‚©‚甉肵‚ؤگ¶’·‚ًٹJژn‚·‚é‚à‚ج‚ئچl‚¦‚ç‚ê‚éپB‚µ‚©‚µپA”‰èچإ“K‰·“x‚ً‰z‚¦‚é‚ئ”‰è—¦‚ھ’ل‚‚ب‚邱‚ئ‚©‚çپAچ‚‰·‚ة‚و‚é“ٌژں‹x–°‚ج‰آ”\گ«‚ھ‚ ‚邱‚ئ‚ھژ¦چ´‚³‚ê‚éپB‚±‚ج‚½‚ك–{ژي‚حپAƒVپ[ƒhƒoƒ“ƒN‚ًŒ`گ¬‚·‚é‰آ”\گ«‚ھ‚ ‚é‚ئچl‚¦‚ç‚ê‚éپB
3.ƒCƒkƒrƒG(ƒCƒl‰بˆê”Nگ¶‘گ–{پAEchinochloa crus-galli var. crus-galli)
پ@–{ژي‚ح“ْ–{‚جچف—ˆژي‚إپA‘Sچ‘‚جژ¼’n‚ةگ¶ˆç‚µ‚ؤ‚¢‚é‚ئ‚³‚ê‚éپi—ر1983پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA‰·“x‚ھچ‚‚‚ب‚é‚ة‚آ‚ê‚ؤچ‚‚‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/6پژ‚إ‚حچإڈI”‰è—¦‚ھ•½‹د‚إ2%‚إ‚ ‚èپA17/8پژپA22/10پژ‚إ‚ح100%‚إ‚ ‚èپA25/13پژ‚إ‚ح94%پA30/15پژ‚ة‚¨‚¢‚ؤ‚ح100%‚ھ”‰è‚µ‚½(•\8پAگ}6پAگ}27)پB
پ@‚±‚ج‚و‚¤‚ب‰·“xˆث‘¶گ«‚ً—L‚·‚éژيژq‚ًگ¶ژY‚·‚éگA•¨‚حپA’n‰·‚ج’ل‚¢“~‹G‚ة‚ح”‰è‚¹‚¸‚ةپAڈt‚ة‚ب‚ء‚ؤ’n‰·‚ھچ‚‚‚ب‚ء‚ؤ‚©‚甉肵‚ؤگ¶’·‚ًٹJژn‚·‚é‚à‚ج‚ئچl‚¦‚ç‚ê‚éپB’ل‰·‚إ”‰è‚ھ‹ة’[‚ة—}گ§‚³‚ê‚邱‚ئ‚©‚çپA–{ژي‚ح‘پڈt‚ج–¾‚é‚¢—§’nپپ‹Gگك“IƒMƒƒƒbƒv‚ً—ک—p‚إ‚«‚ب‚¢ژي‚ئچl‚¦‚ç‚ê‚éپB‚±‚ج‚½‚ك‚ة–{ژي‚حپAک—گ’n‚ب‚ا‚ج—‡’n‚ة‘½‚گ¶ˆç‚µپA—ر“à‚ب‚ا’n‰·‚جڈمڈ¸‚ھ—}گ§‚³‚ê‚é—§’n‚إ‚ح‚ ‚ـ‚èگ¶ˆç‚µ‚ب‚¢‚ئچl‚¦‚ç‚ê‚éپB
4.ƒCƒmƒRƒYƒ`(ƒqƒ†‰ب‘½”Nگ¶‘گ–{پAAchyranthes japonica)
پ@–{ژي‚ح“ْ–{‚جچف—ˆژي‚إپA‘Sچ‘‚جژR’nپA—ر‰؛پA’|‚â‚ش‚ب‚ا‚ةگ¶ˆç‚µ‚ؤ‚¢‚é‚ئ‚³‚ê‚éپi—ر1983پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚ح‚ظ‚ئ‚ٌ‚ا•د‰»‚ھ‚ب‚پA‚·‚ׂؤ‚ج‰·“xڈًŒڈ‚إچ‚‚¢”‰è—¦‚ئ‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/‚Uپژ‚إ‚ح•½‹د‚إ–ٌ99%پA17/8پژ‚إ‚ح98%پA22/10پA25/13پA30/15پژ‚ة‚¨‚¢‚ؤ‚ح100%‚ھ”‰è‚µ‚½(•\8پAگ}7پAگ}25)پB
5.ƒEƒVƒnƒRƒx(ƒiƒfƒVƒR‰ب‰z”Nگ¶‘گ–{پAStellaria aquatica)
پ@–{ژي‚ح“ْ–{‚جچف—ˆژي‚إپA‘Sچ‘‚جژR–ى‚ةگ¶ˆç‚µ‚ؤ‚¢‚é‚ئ‚³‚ê‚éپi—ر1983پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA‰·“x‚ھچ‚‚‚ب‚é‚ة‚آ‚ê‚ؤچ‚‚‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/6پژ‚إ‚حچإڈI”‰è—¦‚ح•½‹د‚إ12%‚¾‚ء‚½‚ج‚ة‘خ‚µ‚ؤپA17/8پژ‚إ‚ح–ٌ79%‚ئ‚ب‚èپA22/10پژ‚إ‚ح88%پA25/13پژ‚إ‚ح94%پA30/15پژ‚ة‚¨‚¢‚ؤ‚ح–ٌ93%‚ھ”‰è‚µ‚½(•\8پAگ}8پAگ}27)پB
6.ƒIƒIƒAƒŒƒ`ƒmƒMƒN(ƒLƒN‰ب‰z”Nگ¶‘گ–{پAErigeron sumaterensis)
پ@–{ژي‚حƒuƒ‰ƒWƒ‹Œ´ژY‚إپA“ْ–{‚ة‚ح‘هگ³––ٹْ‚©‚çگN“ü‹Lک^‚ھ‚ ‚éپBŒ»چفپAٹض“Œˆبگ¼‚جچr‚ê’n‚⓹‚خ‚½‚ب‚ا‚ة’è’…‚µ‚ؤ‚¢‚é‚ئ‚³‚ê‚éپiگ´گ…¥گX“c¥œA“c2001پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA22/10پژ‚إچإ‘ه‚ئ‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/6پژ‚إ‚ح•½‹د‚إ10%پA17/8پژ‚إ‚ح–ٌ11%پA22/10‚إ‚ح–ٌ25%پA25/13‚إ‚ح18%‚إ‚ ‚èپA30/15پژ‚إ‚ح–ٌ9%‚ھ”‰è‚µ‚½(•\8پAگ}9پAگ}26)پB
پ@‚±‚ج‚و‚¤‚ب‰·“xˆث‘¶گ«‚ً—L‚·‚éژيژq‚ًگ¶ژY‚·‚éگA•¨‚حپA’n‰·‚ج’ل‚¢“~‹G‚ة‚ح”‰è‚¹‚¸‚ةپA”سڈt‚ة‚ب‚ء‚ؤ’n‰·‚ھ20پژˆبڈم‚ئچ‚‚‚ب‚ء‚ؤ‚©‚甉肵پAگ¶’·‚ًٹJژn‚·‚é‚ئچl‚¦‚ç‚ê‚éپB‚µ‚©‚µپA”‰èچإ“K‰·“x‚ً‰z‚¦‚é‚ئ”‰è—¦‚ھ’ل‚‚ب‚邱‚ئ‚©‚çپAچ‚‰·‚ة‚و‚é“ٌژں‹x–°‚ج‰آ”\گ«‚ھ‚ ‚邱‚ئ‚ھژ¦چ´‚³‚ê‚éپB‚±‚ج‚½‚ك–{ژي‚حپAƒVپ[ƒhƒoƒ“ƒN‚ًŒ`گ¬‚·‚é‰آ”\گ«‚ھ‚ ‚é‚ئچl‚¦‚ç‚ê‚éپB
7.ƒIƒIƒjƒVƒLƒ\ƒE(ƒgƒEƒ_ƒCƒOƒT‰بˆê”Nگ¶‘گ–{پAEuphorbia maculata)
پ@–{ژي‚ح–kƒAƒپƒٹƒJŒ´ژY‚إپA“ْ–{‚ة‚ح–¾ژ،––ٹْ‚©‚çگN“ü‹Lک^‚ھ‚ ‚éپBŒ»چفپA‘Sچ‘‚ج–q‘گ’nپA”¨’nپA“S“¹‰ˆگüپAچr‚ê’nپA“¹‚خ‚½‚ب‚ا‚ة’è’…‚µ‚ؤ‚¢‚é‚ئ‚³‚ê‚éپiگ´گ…¥گX“c¥œA“c2001پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA‰·“x‚ھچ‚‚‚ب‚é‚ة‚آ‚ê‚ؤچ‚‚‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/6پژ‚إ‚حچإڈI”‰è—¦‚ح•½‹د‚إ10%پA17/8پژ‚إ‚ح–ٌ19%‚إ‚ ‚èپA22/10پژ‚إ‚ح–ٌ43%پA25/13پژ‚إ‚ح–ٌ45%پA30/15پژ‚ة‚¨‚¢‚ؤ‚ح–ٌ53%‚ھ”‰è‚µ‚½(•\8پAگ}10پAگ}27)پB
8.ƒJƒ[ƒNƒT(ƒCƒl‰ب‘½”Nگ¶‘گ–{پAErangrostis ferruginea)
پ@–{ژي‚ح“ْ–{‚جچف—ˆژي‚إپA‘Sچ‘‚ج“¹‚خ‚½‚â‹َ‚«’n‚ب‚ا‚ةگ¶ˆç‚µ‚ؤ‚¢‚éپi—ر1983پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA‰·“x‚ھچ‚‚‚ب‚é‚ة‚آ‚ê‚ؤچ‚‚‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/6پژ‚إ‚ح‘S‚”‰è‚µ‚ب‚©‚ء‚½‚ھپA17/8پژ‚إ‚حچإڈI”‰è—¦‚ح•½‹د‚إ80%‚إ‚ ‚èپA22/10پژ‚إ‚ح–ٌ89%پA25/13پژ‚إ‚ح–ٌ87%پA30/15پژ‚ة‚¨‚¢‚ؤ‚ح96%‚ھ”‰è‚µ‚½(•\8پAگ}11پAگ}27)پB
9.ƒJƒ“ƒgƒEƒˆƒپƒi(ƒLƒN‰ب‘½”Nگ¶‘گ–{پAKalimeris pseudoyomena)
پ@–{ژي‚ح“ْ–{‚جچف—ˆژي‚إپA–{ڈBٹض“Œˆب–k‚ج“c‚ج‚ ‚؛‚âگى‚ׂè‚ب‚ا‚ةگ¶ˆç‚µ‚ؤ‚¢‚éپi—ر1983پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚é25/13پژ‚إ‚جچإڈI”‰è—¦‚حپA0%‚إ‚ ‚ء‚½(•\8پAگ}12)پB
10.ƒMƒVƒMƒV(ƒ^ƒf‰ب‘½”Nگ¶‘گ–{پARumex japonicus)
پ@–{ژي‚ح“ْ–{‚جچف—ˆژي‚إپA‘Sچ‘‚ج“c”¨‚ج‚ ‚؛پAژ¼‚ء‚½“¹‚خ‚½پA–ىŒ´‚ب‚ا‚ةگ¶ˆç‚µ‚ؤ‚¢‚éپi—ر1983پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA‰·“x‚ھچ‚‚‚ب‚é‚ة‚آ‚ê‚ؤچ‚‚‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/6پژ‚إ‚حچإڈI”‰è—¦‚ح•½‹د‚إ–ٌ49%‚إ‚ ‚èپA17/8پژ‚إ‚ح–ٌ96%پA22/10پژ‚إ‚ح–ٌ95%پA25/13پژ‚إ‚ح–ٌ97%پA30/15پژ‚ة‚¨‚¢‚ؤ‚ح–ٌ95%‚ھ”‰è‚µ‚½(•\8پAگ}13پAگ}27)پB10/6پژ‚ة‚¨‚¢‚ؤ–ٌ49%‚ھ”‰è‚µپA‚ـ‚½4پژ‚إ‚ج—âژ¼ڈˆ—ٹْٹش’†‚ة‚à–ٌ83%‚ھ”‰è‚µ‚½پi•\9پA10پjپB‚ـ‚½پA”|—{30“ْŒم‚ةژ{‚µ‚½“ٌ“x–ع‚ج—âژ¼ڈˆ—ٹْٹش’†‚ة‚à‚³‚ç‚ة–ٌ14%‚ھ”‰è‚µ‚½پB
11.ƒMƒjƒAƒOƒ‰ƒX(ƒCƒl‰ب‘½”Nگ¶‘گ–{پAPanicum maximum Jacq.)
پ@–{ژي‚ح“ىƒAƒtƒٹƒJŒ´ژY‚إپA“ْ–{‚ة‚ح1970”N‘م‚©‚çگN“ü‹Lک^‚ھ‚ ‚éپBŒ»چفپAٹض“Œˆبگ¼‚ج”_چk’nژü•س‚ة’è’…‚µ‚ؤ‚¢‚é‚ئ‚³‚ê‚éپiگ´گ…¥گX“c¥œA“c2001پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚ج‚·‚ׂؤ‚ة‚¨‚¢‚ؤچ‚‚¢’l‚ئ‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/‚Uپژ‚إ‚ح–ٌ71%پA17/8پA22/10پA25/13پA30/15پژ‚ة‚¨‚¢‚ؤ‚ح100%‚ھ”‰è‚µ‚½(•\8پAگ}14پAگ}25)پB
پ@‚±‚ج‚و‚¤‚ب‰·“xˆث‘¶گ«‚ً—L‚·‚éژيژq‚ًگ¶ژY‚·‚éگA•¨‚حپA’n‰·‚ج’ل‚¢“~‹G‚ة‚à”‰è‚µپAگ¶’·‚ًٹJژn‚·‚é‚ئچl‚¦‚ç‚ê‚éپB‚ـ‚½ژژŒ±‚µ‚½‘S‰·“xˆو‚إچإڈI”‰è—¦‚ھ90%ˆبڈم‚إ‚ ‚邱‚ئ‚©‚çپA–{ژي‚ح“yڈëƒVپ[ƒhƒoƒ“ƒN‚ًŒ`گ¬‚·‚é‰آ”\گ«‚ح”ٌڈي‚ةڈ‚ب‚¢‚ئگ„ژ@‚³‚ê‚éپB‚·‚ب‚ي‚؟پA2003”N‚و‚èƒrƒIƒgپ[ƒv“à‚إڈoŒ»‚µ‹}‘¬‚ةٹg‘ه‚µ‚ؤ‚¢‚é–{ژي‚ًپAژيژq‚ًگ¶ژY‚·‚é‘O‚ةٹ ‚èژو‚è‚âˆّ‚«”²‚‚±‚ئ‚ة‚و‚èپAƒrƒIƒgپ[ƒv“à‚©‚çڈœ‹ژ‚·‚邱‚ئ‚ھ‰آ”\‚إ‚ ‚é‚ئچl‚¦‚ç‚ê‚éپB
12.ƒRƒZƒ“ƒ_ƒ“ƒOƒT(ƒLƒN‰بˆê”Nگ¶‘گ–{پABidens pilosa L.)
پ@–{ژي‚ح”M‘رƒAƒپƒٹƒJŒ´ژY‚إپA“ْ–{‚ة‚حچ]Œثژ‘م‚©‚çگN“ü‹Lک^‚ھ‚ ‚éپBŒ»چفپA–{ڈB’†•”ˆبگ¼‚ج”¨’nپAژ÷‰€’nپA–q‘گ’nپAژإ’nپA“¹‚خ‚½پAچr‚ê’n‚ب‚ا‚ة’è’…‚µ‚ؤ‚¢‚éپiگ´گ…¥گX“c¥œA“c2001پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA‰·“x‚ھچ‚‚‚ب‚é‚ة‚آ‚ê‚ؤچ‚‚‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/6پژ‚إ‚حچإڈI”‰è—¦‚ھ•½‹د‚إ–ٌ45%‚إ‚ ‚èپA17/8پژ‚إ‚ح94%پA22/10پA25/13پژ‚إ‚ح100%پA30/15پژ‚ة‚¨‚¢‚ؤ‚ح98%‚ھ”‰è‚µ‚½(•\8پAگ}15پAگ}27)پB10/6پژ‚إ–ٌ45%‚ھ”‰è‚µپA‚ـ‚½4پژ‚إ‚ج—âژ¼ڈˆ—ٹْٹش’†‚ة‚à–ٌ99%‚ھ”‰è‚µ‚½پi•\9پA10پjپB‚ـ‚½پA”|—{30“ْŒم‚ةژ{‚µ‚½“ٌ“x–ع‚ج—âژ¼ڈˆ—ٹْٹش’†‚ة‚à‚³‚ç‚ة16%‚ھ”‰è‚µ‚½پB
13.ƒXƒXƒL(ƒCƒl‰ب‘½”Nگ¶‘گ–{پAMiscanthus sinensis)
پ@–{ژي‚ح“ْ–{‚جچف—ˆژي‚إپA‘Sچ‘‚ج•½’n‚âژR’n‚ج“ْ“–‚½‚è‚ج‚و‚¢ڈêڈٹ‚ةگ¶ˆç‚µ‚ؤ‚¢‚éپi—ر1983پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA‰·“x‚ھچ‚‚‚ب‚é‚ة‚آ‚ê‚ؤچ‚‚‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/6پژ‚إ‚ح‘S‚”‰è‚µ‚ب‚©‚ء‚½‚ج‚ة‘خ‚µ‚ؤپA17/8پژ‚إ‚حچإڈI”‰è—¦‚ھ•½‹د‚إ–ٌ33%‚إ‚ ‚èپA22/10پژ‚إ‚ح44%پA25/13پژ‚إ‚ح38%پA30/15پژ‚ة‚¨‚¢‚ؤ‚ح40%‚ھ”‰è‚µ‚½(•\8پAگ}16پAگ}27)پB
14.ƒZƒCƒ^ƒJƒAƒڈƒ_ƒ`ƒ\ƒE(ƒLƒN‰ب‘½”Nگ¶‘گ–{پASolidago altissima)
پ@–{ژي‚ح–kƒAƒپƒٹƒJŒ´ژY‚إپA“ْ–{‚ة‚ح‘و“ٌژںگ¢ٹE‘هگيŒم‚©‚çگN“ü‹Lک^‚ھ‚ ‚éپBŒ»چفپA‘Sچ‘‚ج‹َ‚«’n‚â‰حگى•~‚ة’è’…‚µ‚ؤ‚¢‚é‚ئ‚³‚ê‚éپiگ´گ…¥گX“c¥œA“c2001پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA‘S‘ج‚ئ‚µ‚ؤ”ٌڈي‚ةچ‚‚پA‚³‚ç‚ة25/13پژ‚إچإ‘ه‚ئ‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/‚Uپژ‚إ‚ح•½‹د‚إ–ٌ85%پA17/8پژ‚إ‚ح86%پA22/10‚إ‚ح–ٌ93%پA25/13‚إ‚ح–ٌ99%‚إ‚ ‚èپA30/15پژ‚إ‚ح–ٌ85%‚ھ”‰è‚µ‚½(•\8پAگ}17پAگ}26)پB
پ@‚±‚ج‚و‚¤‚ب‰·“xˆث‘¶گ«‚ً—L‚·‚éژيژq‚ًگ¶ژY‚·‚éگA•¨‚حپA’n‰·‚ج’ل‚¢“~‹G‚ًٹـ‚ك‚ؤˆê”N’†پA“yڈëگ…•ھ‚ھڈ\•ھ‚ة‚ ‚肳‚¦‚·‚ê‚خ”‰è‚µ‚ؤپAگ¶’·‚·‚é‚ئچl‚¦‚ç‚ê‚éپB
15.ƒ`ƒJƒ‰ƒVƒo(ƒCƒl‰ب‘½”Nگ¶‘گ–{پAPennisetum alopecuroides)
پ@–{ژي‚ح“ْ–{‚جچف—ˆژي‚إپA‘Sچ‘‚ج“¹‚خ‚½‚ةگ¶ˆç‚µ‚ؤ‚¢‚éپi—ر1983پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA‰·“x‚ھچ‚‚‚ب‚é‚ة‚آ‚ê‚ؤچ‚‚‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/6پژ‚إ‚حچإڈI”‰è—¦‚ھ•½‹د‚إ–ٌ1%‚إ‚ ‚èپA17/8پA22/10پA25/13پA30/15پژ‚ة‚¨‚¢‚ؤ‚ح100%‚ھ”‰è‚µ‚½(•\8پAگ}18پAگ}27)پB
16.ƒ`ƒaƒ~ƒUƒT(ƒCƒl‰ب‘½”Nگ¶‘گ–{پAOplismenus undulatifolius)
پ@–{ژي‚ح“ْ–{‚جچف—ˆژي‚إپA‘Sچ‘‚جژR–ى‚ج—ر“à‚ةگ¶ˆç‚µ‚ؤ‚¢‚éپi—ر1983پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA‰·“x‚ھچ‚‚‚ب‚é‚ة‚آ‚ê‚ؤچ‚‚‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/6پژ‚إ‚حچإڈI”‰è—¦‚ھ•½‹د‚إ–ٌ1%‚إ‚ ‚èپA17/8پA22/10پA25/13پژ‚إ‚ح100%پA30/15پژ‚ة‚¨‚¢‚ؤ‚ح–ٌ97%‚ھ”‰è‚µ‚½(•\8پAگ}19پAگ}27)پB
17.ƒnƒCƒ`ƒSƒUƒT(ƒCƒl‰ب‘½”Nگ¶‘گ–{پAIsachne nipponensis)
پ@–{ژي‚ح“ْ–{‚جچف—ˆژي‚إپAٹض“Œˆبگ¼‚جژ¼’n‚ةگ¶ˆç‚µ‚ؤ‚¢‚éپi—ر1983پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA‚·‚ׂؤ‚ة‚¨‚¢‚ؤچ‚‚¢’l‚ئ‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/‚Uپژ‚إ‚ح–ٌ99%پA17/8پژ‚إ‚ح–ٌ98%پA22/10پA25/13پA30/15پژ‚ة‚¨‚¢‚ؤ‚ح100%‚ھ”‰è‚µ‚½(•\8پAگ}20پAگ}25)پB‚³‚ç‚ة‚حپA4پژ‚إ‚ج—âژ¼ڈˆ—ٹْٹش’†‚ة‚à–ٌ69%‚ھ”‰è‚µ‚½پi•\9پjپB
پ@‚±‚ج‚و‚¤‚ب‰·“xˆث‘¶گ«‚ً—L‚·‚éژيژq‚ًگ¶ژY‚·‚éگA•¨‚حپA’n‰·‚ج’ل‚¢“~‹G‚ة‚à”‰è‚µپAگ¶’·‚ًٹJژn‚·‚é‚ئچl‚¦‚ç‚ê‚éپB‚ـ‚½چإڈI”‰è—¦‚ھ90%ˆبڈم‚إ‚ ‚邱‚ئ‚©‚çپA–{ژي‚ح“yڈëƒVپ[ƒhƒoƒ“ƒN‚ًŒ`گ¬‚·‚é‰آ”\گ«‚ح”ٌڈي‚ةڈ‚ب‚¢‚ئچl‚¦‚ç‚ê‚éپB
18.ƒwƒ‰ƒIƒIƒoƒR(ƒIƒIƒoƒR‰بˆê”Nگ¶‘گ–{پAPlantago lanceolata)
پ@–{ژي‚حƒˆپ[ƒچƒbƒpŒ´ژY‚إپA“ْ–{‚ة‚حچ]Œثژ‘م––ٹْ‚©‚çگN“ü‹Lک^‚ھ‚ ‚éپBŒ»چفپA‘Sچ‘‚ج“¹‚خ‚½پAچr‚ê’nپA‹َ‚«’n‚ب‚ا‚ة’è’…‚µ‚ؤ‚¢‚é‚ئ‚³‚ê‚éپiگ´گ…¥گX“c¥œA“c2001پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚é25/13پژ‚إ‚جچإڈI”‰è—¦‚حپA56%‚إ‚ ‚ء‚½(•\8پAگ}21)پB‚ـ‚½پA”|—{‘O‚ج—âژ¼ڈˆ—ٹْٹش’†‚ة‚à‘S‘ج‚ج–ٌ14%‚ھ”‰è‚µ‚½پi•\9پjپB‚ب‚¨پA–{ژي‚ة‚آ‚¢‚ؤ‚حپAچ،‰ٌچجژو‚µ‚½ژيژqگ”‚ھڈ‚ب‚©‚ء‚½‚ج‚إپAچ،Œم’اژژ‚ھ•K—v‚إ‚ ‚éپB
19.ƒAƒIƒrƒ†(ƒqƒ†‰بˆê”Nگ¶‘گ–{پAAmaranthus viridis L.)
پ@–{ژي‚ح”M‘رƒAƒپƒٹƒJŒ´ژY‚إپA“ْ–{‚ة‚ح‘هگ³––ٹْ‚©‚çگN“ü‹Lک^‚ھ‚ ‚éپBŒ»چفپA‘Sچ‘‚ج”¨’nپA–q‘گ’nپA“¹‚خ‚½پAچr‚ê’n‚ب‚ا‚ة’è’…‚µ‚ؤ‚¢‚é‚ئ‚³‚ê‚éپiگ´گ…¥گX“c¥œA“c2001پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA25پژˆب‰؛‚إ‚ح”ٌڈي‚ة’ل‚©‚ء‚½‚ج‚ة‘خ‚µ‚ؤپA30/15پژ‚إ‚ح‹ة‚ك‚ؤچ‚‚©‚ء‚½پB‚·‚ب‚ي‚؟پA10/6پژ‚إ‚ح0%پA17/8پژ‚إ‚ح–ٌ1%پA22/10پژ‚إ‚ح2%پA25/13پژ‚إ‚ح14%‚إ‚ ‚ء‚½‚ج‚ة‘خ‚µ‚ؤپA30/15پژ‚ة‚¨‚¢‚ؤ‚ح–ٌ91%‚ھ”‰è‚µ‚½(•\8پAگ}22پAگ}28)پB
پ@ˆبڈم‚جŒ‹‰ت‚©‚çپA–{ژي‚حٹh—گ’n‚ب‚ا‚ج—‡’n‚ة‚¨‚¢‚ؤپA’¼ژث“ْŒُ‚ھ“–‚½‚ء‚ؤ’n‰·‚ھ‹ة’[‚ةڈمڈ¸‚·‚邱‚ئ‚ًٹآ‹«ƒVƒOƒiƒ‹‚ئ‚µ‚ؤژَ‚¯‚ؤ”‰è‚µ‚ؤ‚‚éپA“TŒ^“I‚بƒMƒƒƒbƒvŒںڈo‹@چ\‚ً—L‚·‚éژي‚إ‚ ‚é‚ئچl‚¦‚ç‚ê‚éپB–{ƒrƒIƒgپ[ƒv‚ة‚¨‚¢‚ؤ‚حپAڈvچH’¼Œم‚ج2001”N‚ة‚ح‘½گ”‚جƒAƒIƒrƒ†Œآ‘ج‚جگ¶ˆç‚ھٹm”F‚³‚ê‚ؤ‚¢‚½پiگV‰ھ 2002پjپB‚±‚ê‚حپA–{ژي‚ھڈمڈq‚ج‚و‚¤‚بگ¶‘شٹw“I“ءگ«‚ً—L‚µ‚ؤ‚¢‚邱‚ئ‚ة‹Nˆِ‚·‚é‚ئچl‚¦‚ç‚ê‚éپB2003”N“x‚ئچ،‰ٌ‚جگA•¨‘ٹ’²چ¸‚¨‚و‚ر“yڈëƒVپ[ƒhƒoƒ“ƒN‰ًگح‚ة‚¨‚¢‚ؤ‚حپA–{ژي‚ح‚ظ‚ئ‚ٌ‚اŒںڈo‚³‚ê‚ب‚©‚ء‚½‚ج‚إپA–{ƒrƒIƒgپ[ƒv‚ة‚¨‚¢‚ؤ‚ح“yڈëƒVپ[ƒhƒoƒ“ƒN‚ًŒ`گ¬‚µ‚ؤ‚¢‚ب‚¢‚©پA‚µ‚ؤ‚¢‚½‚ئ‚µ‚ؤ‚à”ٌڈي‚ةڈ¬‚³‚ب‚à‚ج‚ة‚ب‚ء‚ؤ‚¢‚é‚ئگ„ژ@‚³‚ê‚éپB
20.ƒˆƒV(ƒCƒl‰ب‘½”Nگ¶‘گ–{پAPhragmites communis)
21.ƒˆƒ‚ƒM(ƒLƒN‰ب‘½”Nگ¶‘گ–{پAArtemisia princeps)
پ@–{ژي‚ح“ْ–{‚جچف—ˆژي‚إپA‘Sچ‘‚جژR’n‚â–ىŒ´‚ب‚ا‚ةگ¶ˆç‚µ‚ؤ‚¢‚éپi—ر1983پjپB–{ژي‚ج”|—{30“ْŒم‚ة‚¨‚¯‚éچإڈI”‰è—¦‚حپAگف’肵‚½‰·“xڈًŒڈ‚ج”حˆح“à‚إ‚حپA‰·“x‚ھچ‚‚‚ب‚é‚ة‚آ‚ê‚ؤچ‚‚‚ب‚èپA22/10پژ‚إچإ‘ه‚ئ‚ب‚ء‚½پB‚·‚ب‚ي‚؟پA10/‚Uپژ‚إ‚ح•½‹د‚إ–ٌ37%پA17/8پژ‚إ‚ح–ٌ43%پA22/10‚إ‚ح–ٌ59%پA25/13‚إ‚ح49%‚إ‚ ‚èپA30/15پژ‚إ‚ح–ٌ55%‚ھ”‰è‚µ‚½(•\8پAگ}24پAگ}26)پB
پ@پ@‚±‚ج‚و‚¤‚ب‰·“xˆث‘¶گ«‚ً—L‚·‚éژيژq‚ًگ¶ژY‚·‚éگA•¨‚حپA’n‰·‚ج’ل‚¢“~‹G‚ًٹـ‚ك‚ؤˆê”N’†پA“yڈëگ…•ھ‚ھڈ\•ھ‚ة‚ ‚肳‚¦‚·‚ê‚خ”‰è‚µ‚ؤپAگ¶’·‚·‚é‚ئچl‚¦‚ç‚ê‚éپB‚µ‚©‚µپA”‰èچإ“K‰·“x‚ً‰z‚¦‚é‚ئ”‰è—¦‚ھ’ل‚‚ب‚邱‚ئ‚©‚çپAچ‚‰·‚ة‚و‚é“ٌژں‹x–°‚ج‰آ”\گ«‚ھ‚ ‚邱‚ئ‚ھژ¦چ´‚³‚ê‚éپB‚±‚ج‚½‚ك–{ژي‚حپAƒVپ[ƒhƒoƒ“ƒN‚ًŒ`گ¬‚·‚é‰آ”\گ«‚à‚ ‚é‚ئچl‚¦‚ç‚ê‚éپB
پ@ˆبڈم21ژي—ق‚جگA•¨‚جژيژq”‰è‚ج‰·“xˆث‘¶گ«‚ًپAچإڈI”‰è—¦‚ً‚à‚ئ‚ة•ھ—ق‚·‚é‚ئپA4‚آ‚جƒOƒ‹پ[ƒv‚ة‘ه•ت‚·‚邱‚ئ‚ھ‚إ‚«‚éپiگ}25پ`28پjپB
پ@ƒOƒ‹پ[ƒv‚`پiگ}25پjپFگف’艷“x”حˆحپi10/6پژپ`30/15پژپj‚ة‚¨‚¢‚ؤ‚حپA‚·‚ׂؤ‚ج‰·“x‹و‚ة‚¨‚¢‚ؤچ‚‚¢پi70پ“ˆبڈمپj”‰è—¦‚ً—L‚·‚éپA‚ـ‚½‚ح‰·“x‹وٹش‚إچإڈI”‰è—¦‚ةچ·‚ھ‚ب‚¢ژيپBƒCƒmƒRƒYƒ`پAƒnƒCƒ`ƒSƒUƒTپAƒˆƒV‚ج3ژي‚جچف—ˆژي‚ئٹO—ˆژي‚جƒMƒjƒAƒOƒ‰ƒX‚ھٹY“–‚·‚éپB‹Gگك‚ً–â‚ي‚¸پA‰·“xˆبٹO‚جٹآ‹«—vˆِپi—ل‚¦‚خ“yڈëگ…•ھپj‚ھ–‚½‚³‚ê‚ê‚خ”‰è‚·‚é‚ئگ„ژ@‚³‚ê‚éپB‚ـ‚½پA“yڈëƒVپ[ƒhƒoƒ“ƒN‚ًŒ`گ¬‚·‚é‰آ”\گ«‚ح’ل‚¢‚ئگ„ژ@‚³‚ê‚éپB
پ@ƒOƒ‹پ[ƒv‚aپiگ}26پjپFگف’艷“x”حˆحپi10/6پژپ`30/15پژپj“à‚إ”‰è‚ةچإ“K‚ب‰·“x‹و‚ھ‘¶چف‚·‚éژيپBچإ“K‰·“x‹و‚حپA22/10پژ‚ـ‚½‚ح25/13پژ‚ئ”نٹr“Iچ‚‚¢پBƒZƒCƒ^ƒJƒAƒڈƒ_ƒ`ƒ\ƒEپAƒCƒgƒoƒMƒNپAƒIƒIƒAƒŒƒ`ƒmƒMƒNپAƒˆƒ‚ƒM‚ھٹY“–‚µپA‚·‚ׂؤƒLƒN‰ب‚جگA•¨‚إ‚ ‚éپBƒˆƒ‚ƒM‚ھچف—ˆژي‚إپA‚»‚êˆبٹO‚ج3ژي‚حٹO—ˆژي‚إ‚ ‚éپBچ‚‰·پi30/15پژپj‚إچإڈI”‰è—¦‚ھ’ل‰؛‚·‚邱‚ئ‚©‚çپAچ‚‰·‚ة‚و‚ء‚ؤ“ٌژں‹x–°‚ھ—U“±‚³‚êپA“yڈëƒVپ[ƒhƒoƒ“ƒN‚ًŒ`گ¬‚·‚é‰آ”\گ«‚ھ‚ ‚é‚ئگ„ژ@‚³‚ê‚éپB
پ@ƒOƒ‹پ[ƒv‚bپiگ}27پjپFگف’艷“x”حˆحپi10/6پژپ`30/15پژپj‚إ‚حپA‰·“x‚ھچ‚‚‚ب‚é‚ة‚آ‚ê‚ؤچإڈI”‰è‚ھچ‚‚‚ب‚éژيپB10/6پژ‚إ‚حچإڈI”‰è—¦‚ھ‘¼‚ج‰·“x‚ئ”ن‚ׂؤ”ٌڈي‚ة’ل‚¢‚±‚ئ‚©‚çپAژه‚ئ‚µ‚ؤڈt‚ة‚ب‚ء‚ؤ’n‰·‚ھ17پژˆبڈم‚ة‚ب‚ء‚ؤ‚©‚çپA”‰è‚·‚é‚ئگ„ژ@‚³‚ê‚éپBƒCƒkƒrƒGپAƒ`ƒaƒ~ƒUƒTپAƒ`ƒJƒ‰ƒVƒoپAƒJƒ[ƒNƒTپAƒMƒVƒMƒVپAƒEƒVƒnƒRƒxپAƒRƒZƒ“ƒ_ƒ“ƒOƒTپAƒAƒپƒٹƒJƒCƒkƒzƒIƒYƒL‚ج8ژي‚ھ‘®‚·ƒTƒuƒOƒ‹پ[ƒv‚إ‚حپA10/6پژˆبٹO‚ج‰·“x‹و‚إ‚ح70پ“ˆبڈم‚ھ”‰è‚·‚éپB‚µ‚½‚ھ‚ء‚ؤ‚±‚ê‚ç‚جژي‚ھ“yڈëƒVپ[ƒhƒoƒ“ƒN‚ًŒ`گ¬‚·‚é‰آ”\گ«‚ح’ل‚¢‚ئگ„ژ@‚³‚ê‚éپB‚ـ‚½ƒIƒIƒjƒVƒLƒ\ƒEپAƒXƒXƒL‚ج2ژي‚ھ‘®‚·‚éƒTƒuƒOƒ‹پ[ƒv‚إ‚حپAچإڈI”‰è—¦‚حچإ‘ه‚إ‚à50پ“’ِ“x‚إ‚ ‚邱‚ئ‚©‚çپAچ‚‰·‚ة‚و‚ء‚ؤ“ٌژں‹x–°‚ھ—U“±‚³‚êپA“yڈëƒVپ[ƒhƒoƒ“ƒN‚ًŒ`گ¬‚·‚é‰آ”\گ«‚ھ‚ ‚é‚ئگ„ژ@‚³‚ê‚éپB‚±‚جƒOƒ‹پ[ƒv‚b‚ة‘®‚·ژي‚ج‚¤‚؟چف—ˆژي‚ة‚حƒCƒl‰ب‚ھ‘½‚پi5ژيپjپA‚ـ‚½ٹO—ˆژي‚ح3ژي‚ج‚ف‚إ‚ ‚éپB
پ@ƒOƒ‹پ[ƒv‚cپiگ}28پjپFٹO—ˆژي‚جƒAƒIƒrƒ†‚ج‚ف‚ھ‘®‚·ƒOƒ‹پ[ƒv‚إ‚ ‚éپB–{ژي‚¾‚¯•تƒOƒ‹پ[ƒv‚ة—ق•ت‚µ‚½——R‚حپA–{ژي‚ھ30/15پژ‚إ‚ج‚فچ‚‚¢پi90پ“’ِ“xپj”‰è—¦‚ً—L‚·‚é‚©‚ç‚إ‚ ‚éپB‘Oڈq‚ج‚و‚¤‚ةپA‚±‚ê‚حƒMƒƒƒbƒvŒںڈo‹@چ\‚ئ‚³‚ê‚éپB
پ@ˆبڈم‚ج‚و‚¤‚ةپA”‰è‚ج‰·“xˆث‘¶گ«‚ًƒOƒ‹پ[ƒv‰»‚µ‚ؤ‚àپA“¯ˆêƒOƒ‹پ[ƒv“à‚ة‚حچف—ˆژي‚ئٹO—ˆژي‚ھچ¬چف‚·‚éپB‚µ‚½‚ھ‚ء‚ؤپAƒrƒIƒgپ[ƒv“à‚ةگ¶ˆç‚·‚éگA•¨‚ج”‰è‚ج‰·“xˆث‘¶گ«‚ةٹض‚µ‚ؤپAٹO—ˆژي‚ئچف—ˆژي‚جٹش‚ة–¾ٹm‚بچ·ˆظ‚ح”F‚ك‚ç‚ê‚ب‚¢‚ئ‚¢‚¦‚éپB‚·‚ب‚ي‚؟پA–ىٹO‚إ‚ج”‰è‹Gگك‚ةٹض‚µ‚ؤپAٹO—ˆژي‚ئچف—ˆژي‚جٹش‚ة–¾ٹm‚بچ·ˆظ‚ح‚ب‚پA‚¢‚¸‚ê‚جژگك‚ة‚¨‚¢‚ؤ‚àپAٹO—ˆژي‚ئچف—ˆژي‚ج‘o•û‚ھ”‰è‚µ‚ؤ‚¢‚é‚ئگ„ژ@‚³‚ê‚éپB
ڈtگA•¨‚جٹJ‰شƒtƒFƒmƒچƒWپ[‚ئگA•¨‘ٹ
پ@2004”N“x‚جŒv7‰ٌ‚جŒ»’n’²چ¸‚ة‚و‚èپAƒrƒIƒgپ[ƒv“à‘Sˆو‚ة‚¨‚¢‚ؤ49ژيپiٹO—ˆگA•¨20ژيپAچف—ˆژي29ژيپj‚جگA•¨‚جگ¶ˆç‚ھٹm”F‚³‚ꂽپi•\11پjپB‚ـ‚½پAچ،”N“xٹm”F‚³‚ꂽ49ژي’†پAڈ‰‚ك‚ؤڈoŒ»‚ھٹm”F‚³‚ꂽژي‚ح14ژيپiٹO—ˆگA•¨6ژيپAچف—ˆژي8ژيپj‚إ‚ ‚ء‚½پB
2004”N‚جٹeŒژ‚ةٹm”F‚³‚ꂽٹJ‰شگA•¨ژيگ”‚حپA2Œژ‚ة4ژيپiٹO—ˆژي3ژيپAچف—ˆژي1ژيپjپA3Œژ‚ة5ژيپiٹO—ˆژي3ژيپAچف—ˆژي2ژيپjپA4Œژ‚ة17ژيپiٹO—ˆژي3ژيپAچف—ˆژي14ژيپjپA5Œژ‚ة13ژيپiٹO—ˆژي7ژيپAچف—ˆژي6ژيپjپA6Œژ‚ة4ژيپiٹO—ˆژي2ژيپAچف—ˆژي2ژيپj‚إ‚ ‚ء‚½پiگ}29پjپB‚·‚ب‚ي‚؟پA4ŒژˆبٹO‚جŒژ‚إ‚حپAٹJ‰ش‚ھٹm”F‚³‚ꂽگA•¨‚ج”¼گ”ˆبڈم‚ھٹO—ˆژي‚إ‚ ‚ء‚½پBˆبڈم‚و‚èپAƒrƒIƒgپ[ƒv–{—ˆ‚ج–ع“I‚ة‰ˆ‚ء‚½چإ‚à—اچD‚بŒiٹد‚ح4Œژ‚ة‚ف‚¤‚¯‚ç‚ê‚é‚ئ‚¢‚¦‚éپB
ڈtگA•¨‚ج•ھ•zˆت’uپE•ھ•z–تگد
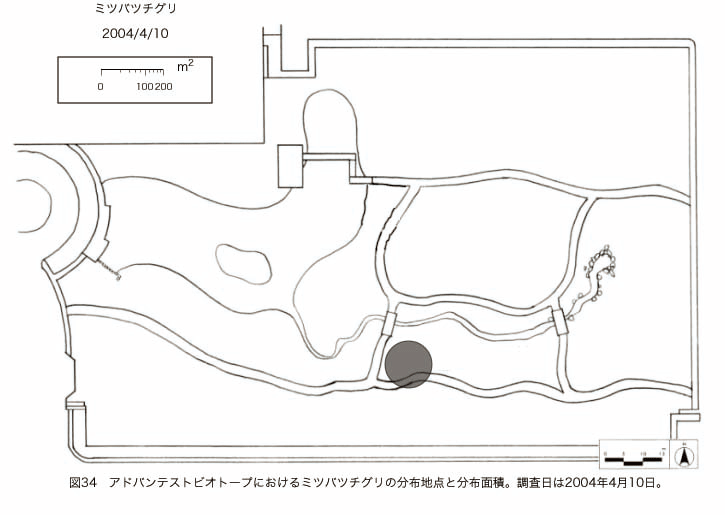
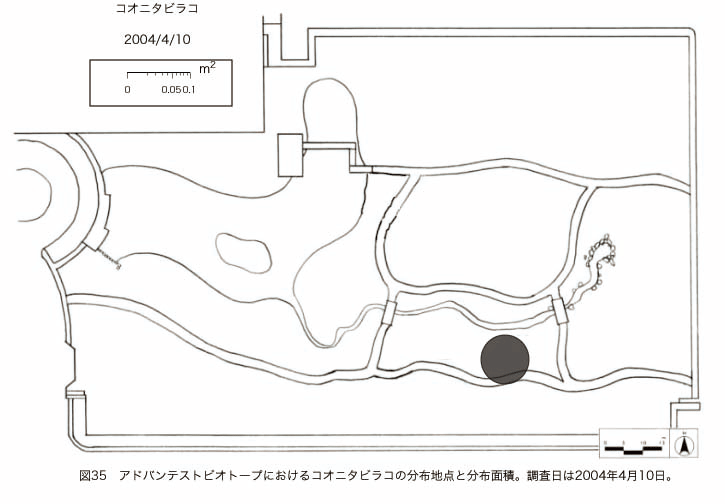
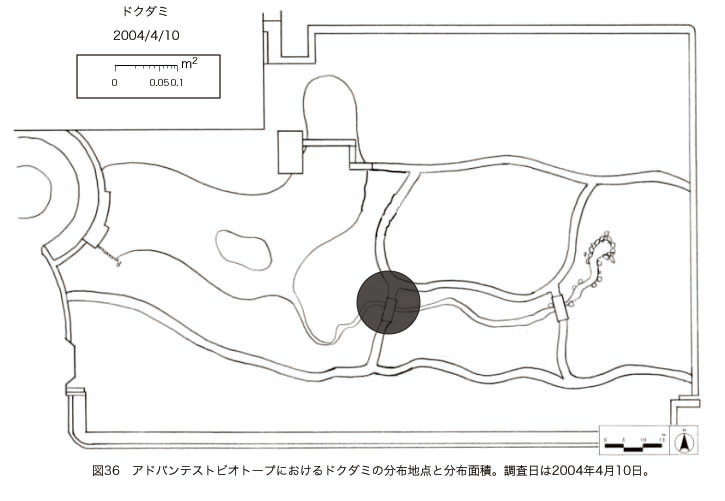
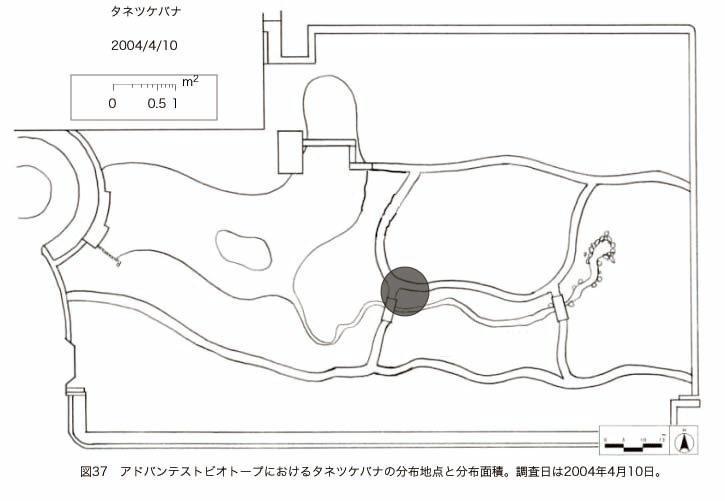
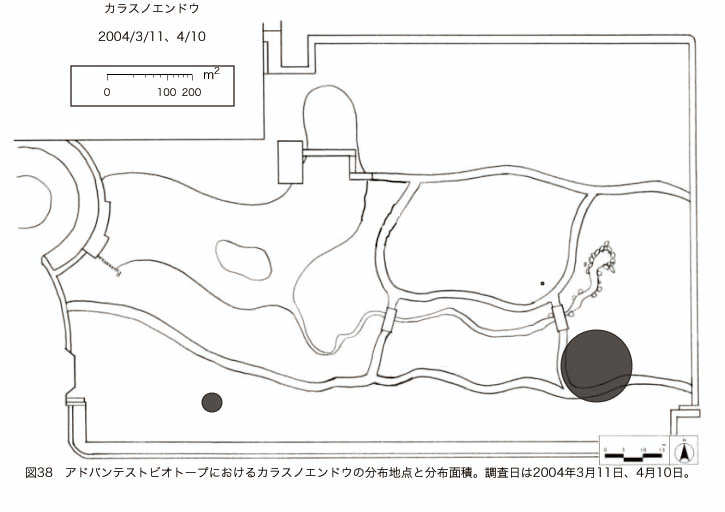
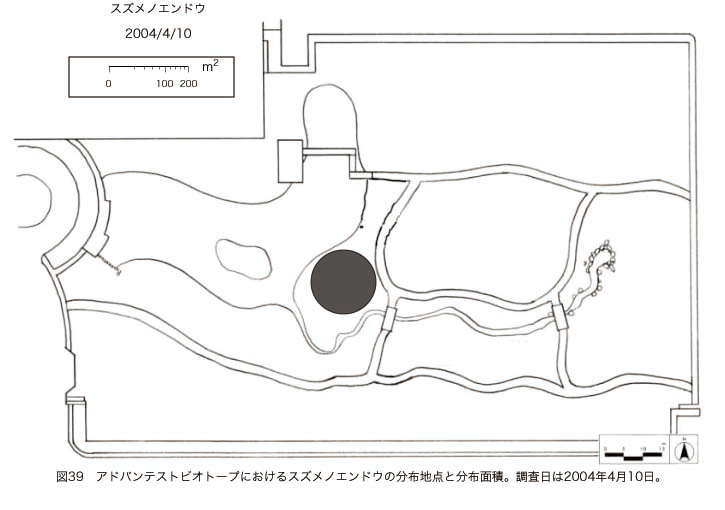
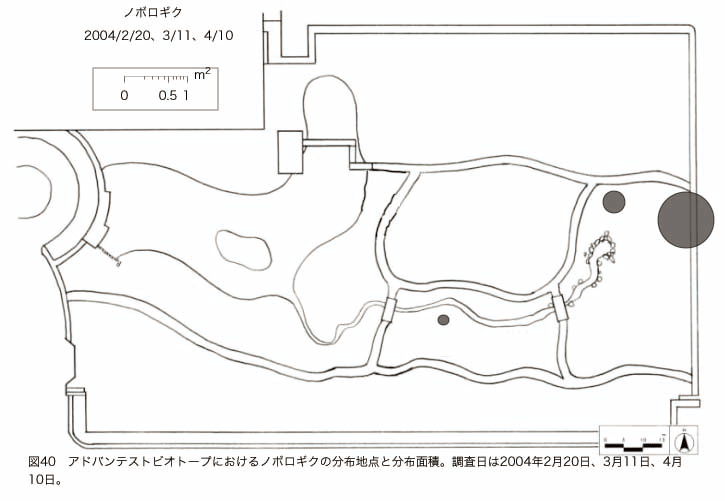
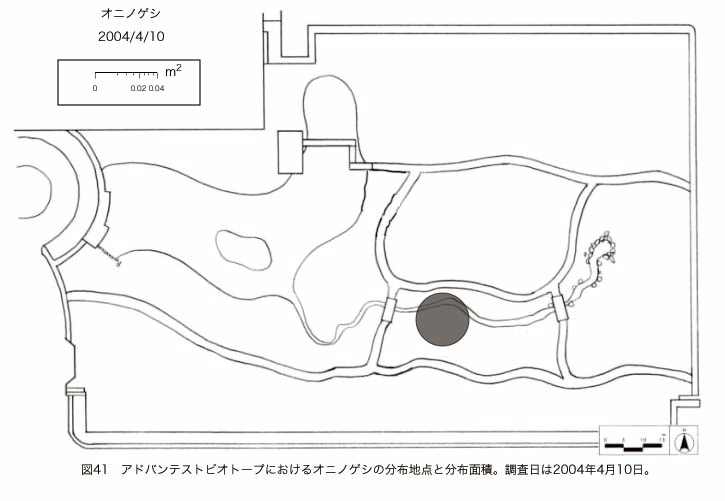
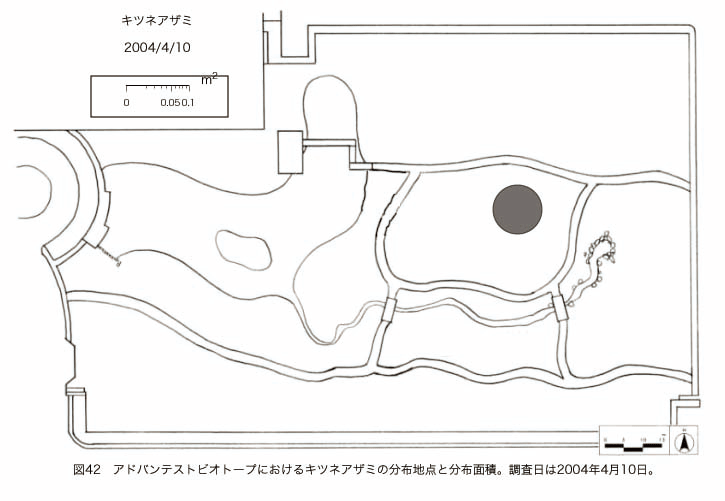
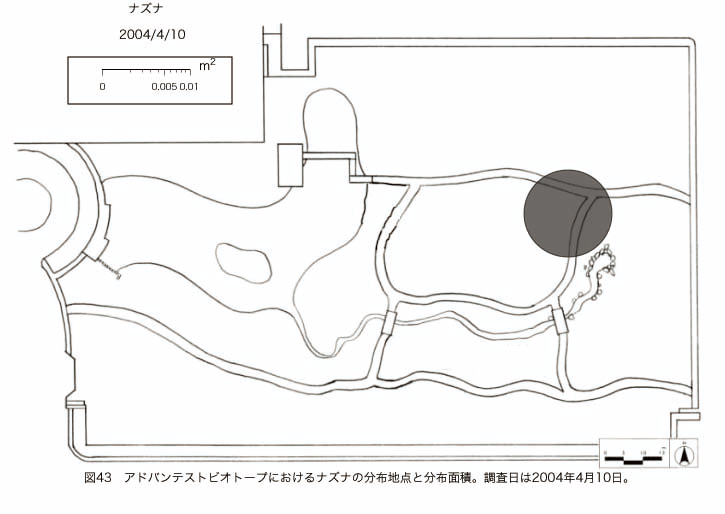
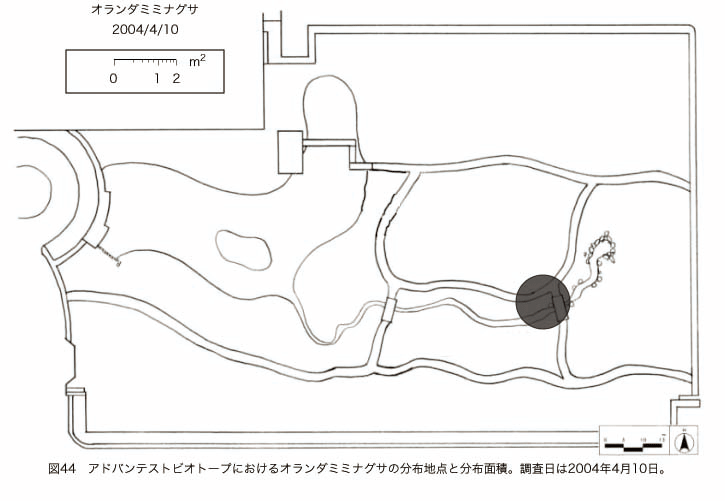
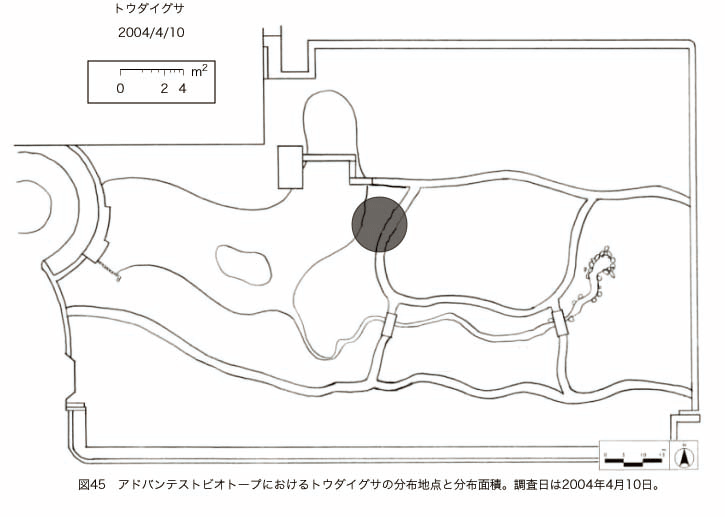
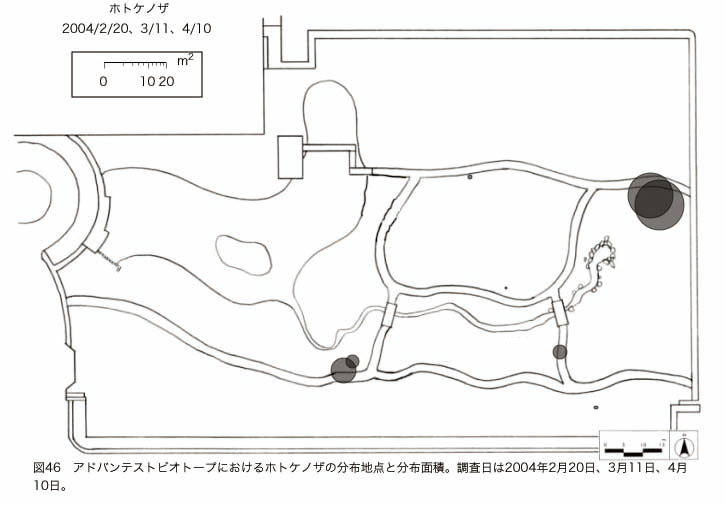
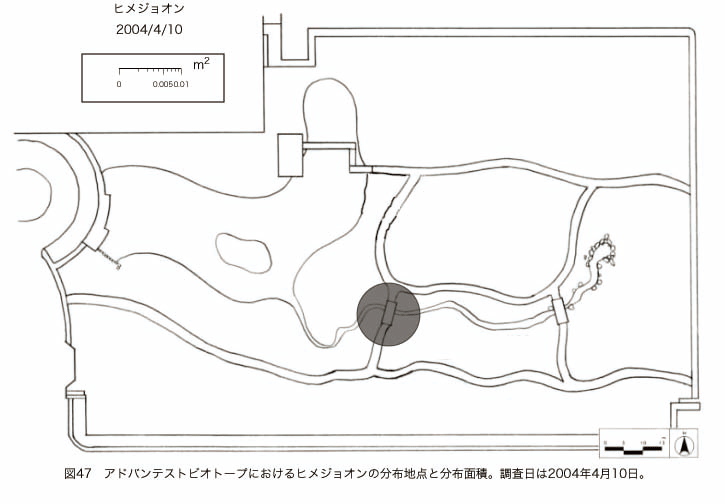
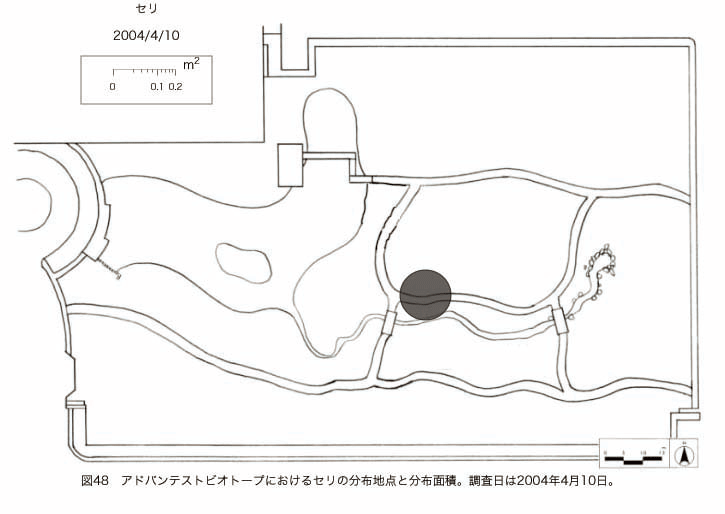
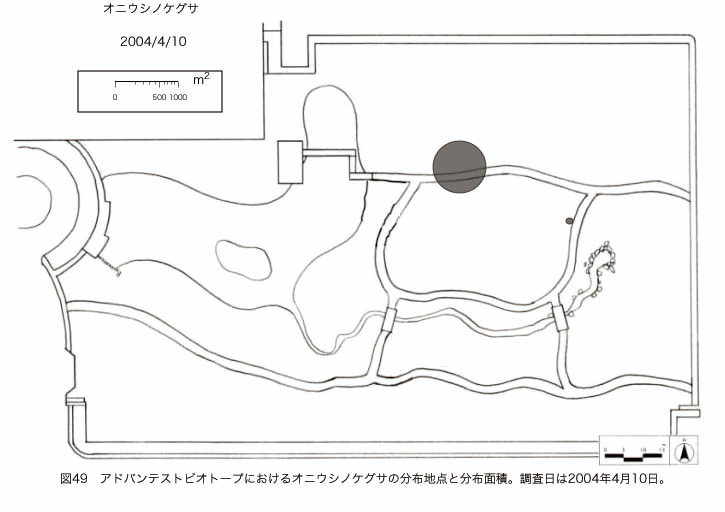
پ@ƒrƒIƒgپ[ƒv“à‚ة‚¨‚¢‚ؤ20ژيپAŒv72’n“_‚جڈtگA•¨‚ھٹm”F‚³‚ꂽپi•\12پjپiگ}29پAگ}30پAگ}31پAگ}32پAگ}33پAگ}34پAگ}35پAگ}36پAگ}37پAگ}38پAگ}39پAگ}40پAگ}41پAگ}42پAگ}43پAگ}44پAگ}45پAگ}46پAگ}47پAگ}48پAگ}49پjپB‚±‚ج‚¤‚؟ٹO—ˆژي‚حƒIƒIƒCƒkƒmƒtƒOƒٹپAƒZƒCƒˆƒEƒ^ƒ“ƒ|ƒ|پAƒIƒ‰ƒ“ƒ_ƒ~ƒ~ƒiƒOƒTپAƒmƒ{ƒچƒMƒNپAƒqƒپƒWƒ‡ƒIƒ“پAƒIƒjƒEƒVƒmƒPƒOƒT‚ج6ژي‚إ‚ ‚ء‚½پBٹm”F‚³‚ꂽ’n“_گ”‚ھچإ‚à‘½‚پA•ھ•z–تگد‚àچإ‘ه‚إ‚ ‚ء‚½‚ج‚حپAٹO—ˆژي‚إ‚ ‚éƒIƒIƒCƒkƒmƒtƒOƒٹپiŒv32’n“_پA–ٌ3910†Fپj‚إ‚ ‚ء‚½پBƒIƒIƒCƒkƒmƒtƒOƒٹ‚حڈtگA•¨پپ“~‹Gˆê”Nگ¶گA•¨‚إپA‘پڈt‚ج‹َ‚«ƒjƒbƒ`‚ً—ک—p‚·‚éƒ^ƒCƒv‚جگA•¨‚إ‚ ‚éپB‚µ‚½‚ھ‚ء‚ؤپA–{ژي‚حٹO—ˆژي‚إ‚ح‚ ‚é‚ھپA”ة–خ‚µ‚ؤ‚àŒم‘±‚·‚éگA•¨‚ض‚ج‰e‹؟‚حڈ¬‚³‚¢‚ج‚إ‚ح‚ب‚¢‚©‚ئگ„ژ@‚³‚ê‚éپBچ،ŒمپAŒم‘±‚·‚éگA•¨‚ة‘خ‚·‚é‰e‹؟‚ھŒںڈo‚³‚ꂽڈêچ‡‚ة‚حڈœ‹ژ‚ًچs‚¤•K—v‚ھ‚ ‚é‚ھپA‚»‚ê‚ـ‚إ‚ح“––تڈœ‹ژ‚ج•K—v‚ح‚ب‚¢‚ئچl‚¦‚ç‚ê‚éپB
ƒMƒjƒAƒOƒ‰ƒX‚ج‘Sچ‘•ھ•z
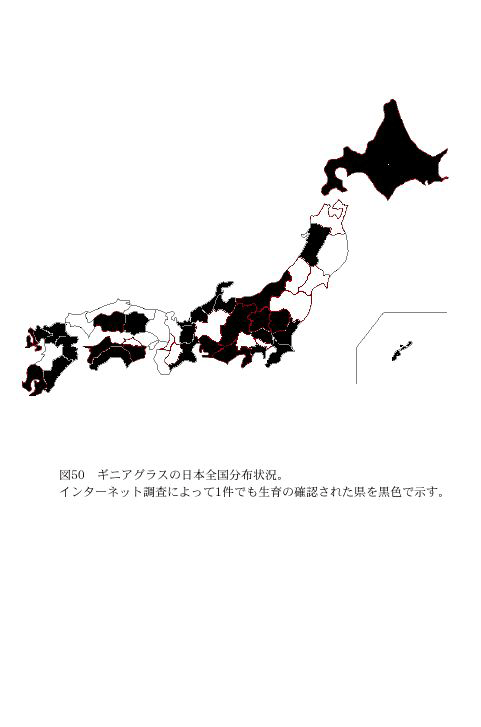
پ@پuYahoo!Japanپv‚ً—p‚¢‚ؤپuƒMƒjƒAƒOƒ‰ƒXپv‚ًƒLپ[ƒڈپ[ƒh‚ئ‚µ‚ؤŒںچُ‚µ‚½Œ‹‰تپA304Œڈ‚جƒzپ[ƒ€ƒyپ[ƒW‚ًŒںڈo‚µ‚½پi2004”N6Œژ30“ْŒ»چفپjپB‚»‚جŒ‹‰تپA“ْ–{‘Sچ‘28Œ§پiگ}50پj‚إگ¶ˆç‚ھٹm”F‚³‚ꂽپBƒMƒjƒAƒOƒ‰ƒX‚حژ”—؟—p‚ئ‚µ‚ؤ‚¾‚¯‚إ‚ح‚ب‚پAچى•¨‚جکAچىڈلٹQ‚âƒZƒ“ƒ`ƒ…ƒE–hڈœ‚ج‚½‚ك‚ة”¨’n‚إچح”|‚³‚ê‚ؤ‚¢‚邱‚ئ–¾‚ç‚©‚ة‚ب‚ء‚½پB‚·‚ب‚ي‚؟ژg—p—p“r‚حپAژ”—؟—p16Œ§پAکAچىڈلٹQپEƒZƒ“ƒ`ƒ…ƒE–hڈœ—p13Œ§‚إ‚ ‚ء‚½پi•\13پjپB‚±‚ج‚و‚¤‚ةٹe’n‚إچح”|‚³‚ê‚ؤ‚¢‚邱‚ئ‚©‚çپAچ،Œم‚à”¨’n‚©‚ç”ٍژU‚µ‚½ƒMƒjƒAƒOƒ‰ƒX‚جژيژq‚ة‚و‚èپA•ھ•z‚ھٹg‘ه‚µ‚ؤ‚¢‚‰آ”\گ«‚ھ‚ ‚é‚ئچl‚¦‚ç‚ê‚éپB
—ر“à‚ج‘ٹ‘خŒُ—تژq–§“x
—ر’²چ¸’n“_‚ج“àŒv16’n“_‚ة‚¨‚¢‚ؤپA‘ٹ‘خŒُ—تژq–§“x‚ح11.6~41.9%‚ج”حˆح‚إ‚ ‚ء‚½پi•\14پjپBŒُ—تژq–§“x10%‚«‚´‚ف‚ج•p“x•ھ•z(گ}51b)‚ة‚¨‚¢‚ؤ‚حپA10~20%‹و•ھ“à‚ة8’n“_‚ھ‚ ‚èپA20~30%‚إ‚ح6’n“_‚ھ‚ ‚ء‚½پB2003”N“x‚ج’²چ¸Œ‹‰ت(گ¯–ى‚ة‚و‚éپAگ}51aپA’²چ¸’n“_گ”‚ح36)‚ئچ،”N“x‚ج’²چ¸Œ‹‰ت‚ً”نٹr‚·‚é‚ئپA2003”N“x‚ة‚¨‚¢‚ؤ‚حپA50%ˆبڈم‚ج’n“_‚ح7’n“_‚إ‚ ‚ء‚½‚ج‚ة‘خ‚µپAچ،”N“x‚ح0’n“_‚ئ‘ه•‚ةŒ¸‚ء‚ؤ‚¢‚½پB‚±‚¤‚µ‚½چ‚‚¢Œُ—تژq–§“x‚ج’n“_‚ج‘ه•‚بŒ¸ڈ‚حپAƒrƒIƒgپ[ƒv‘¢گ¬ژ‚ةگAچح‚³‚ꂽچ‚–ط‚جگ¬’·‚ة‚و‚é‚à‚ج‚ئچl‚¦‚ç‚ê‚éپBٹO—ˆگA•¨‚ج‘½‚‚ح’¼ژثŒُ‚ج‘½‚¢—§’n‚ًچD‚ق‚½‚كپAچ،ŒمƒrƒIƒgپ[ƒv‚ج—ر“à‚ة‚¨‚¢‚ؤٹO—ˆگA•¨‚حŒ¸ڈ‚·‚邱‚ئ‚ھٹْ‘ز‚³‚ê‚éپB
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}