考 察
実験€ 伊勢崎・水上両地点に生育するオオブタクサの生長・種子生産比較
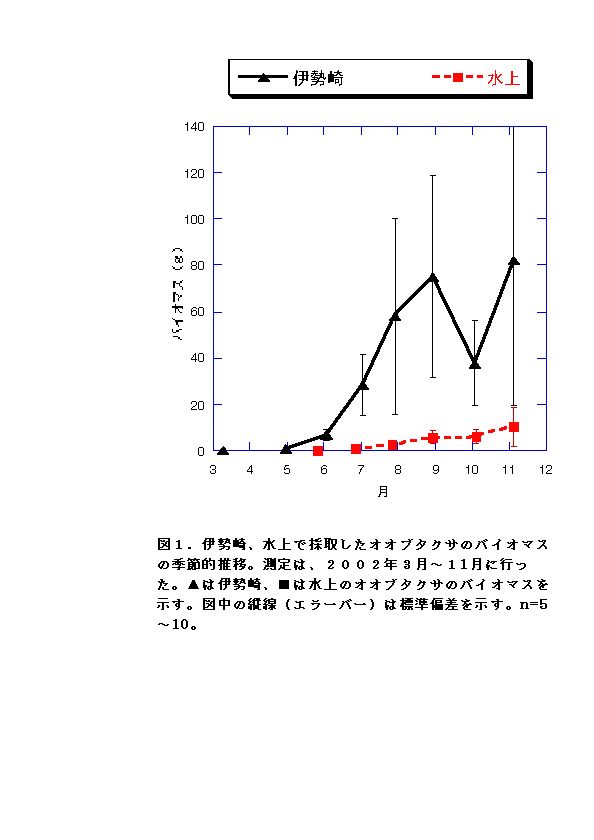
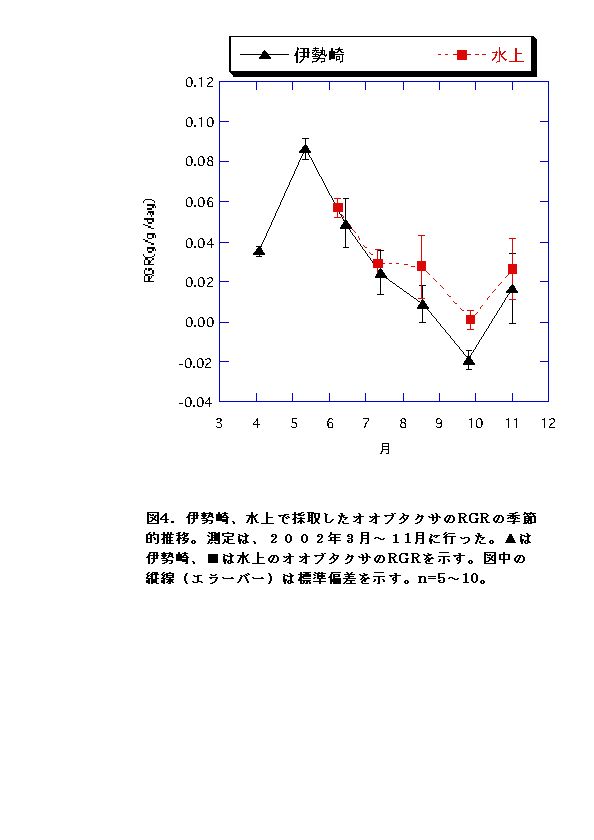
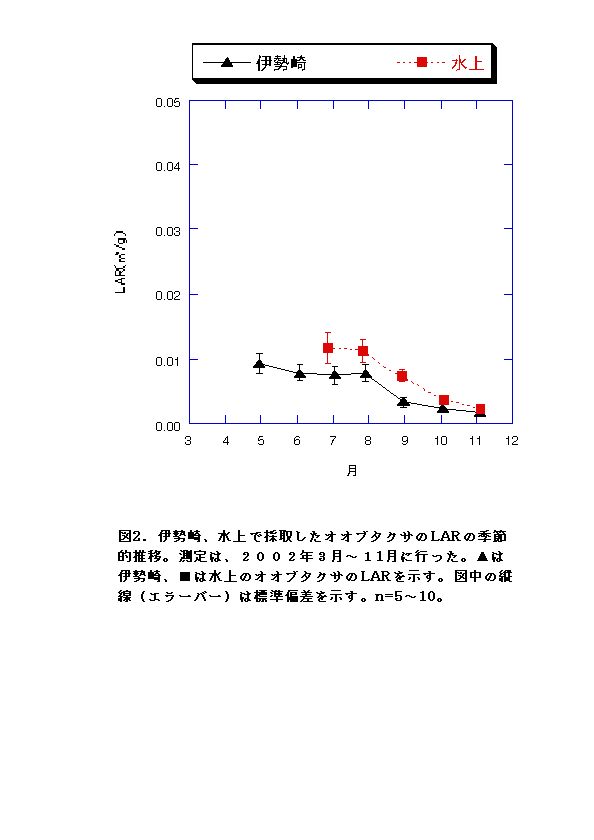
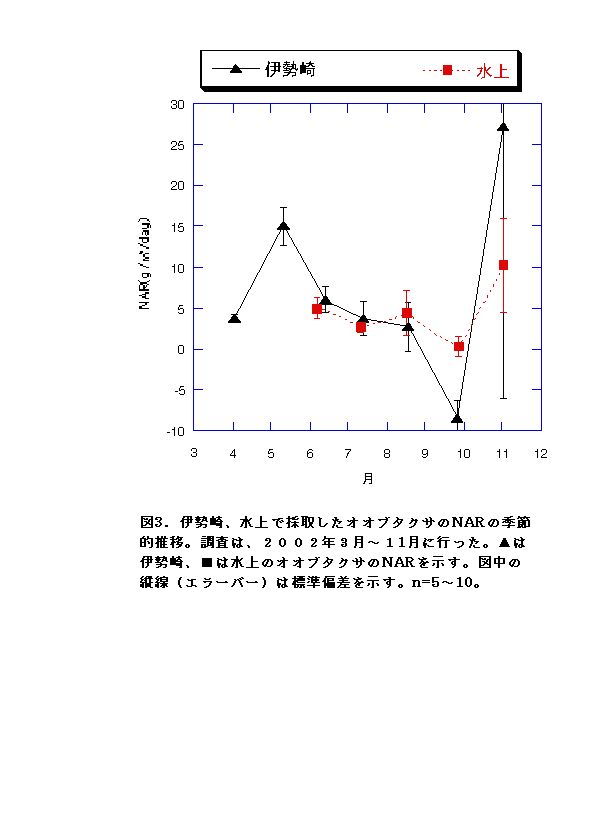
現地個体のバイオマスを比較すると、伊勢崎の調査地のオオブタクサの方が大きい(図1)。しかしRGRはむしろ水上の個体の方が大きくなる月もあった(図4)。このことから、水上のオオブタクサよりも、伊勢崎のオオブタクサの方が現地での生長が速いいうことはないといえる。したがって伊勢崎のオオブタクサのバイオマスが大きくなる原因は、生育期間が長いこと、つまり発芽時期が早いことが考えられる。実際、伊勢崎の調査地のオオブタクサは水上の調査地のオオブタクサよりも2ヶ月も早く発芽していたことが示されている(吉井、2002)。今回の調査でも、伊勢崎の方が水上よりも2ヶ月早く発芽していた。 LARは常に水上の方が高いことと(図2)、NARは両地で顕著な差は見られないこと(図3)から、水上でもRGRが伊勢崎と同程度かそれ以上になる時期がある原因として、葉面積比を高く維持していることが考えられる。
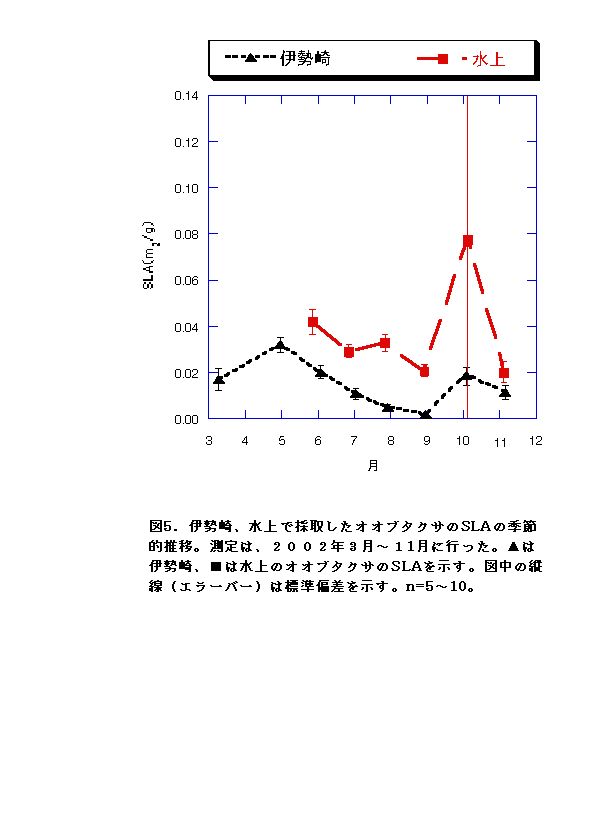
器官別重量比の季節変化傾向には、水上と伊勢崎のオオブタクサで共通性が高い。すなわち、両地共に葉や根の重量は時間とともに減る、もしくは変化がほとんど見られないが、茎の重量は最後まで増え続けた(図5〜8)。このことはオオブタクサの競争者としての特性を強く表していると言える。茎に多くのバイオマスを費やしておけば、より高い位置に葉を展開することができ、生存に不可欠な太陽光を確保できるからである。
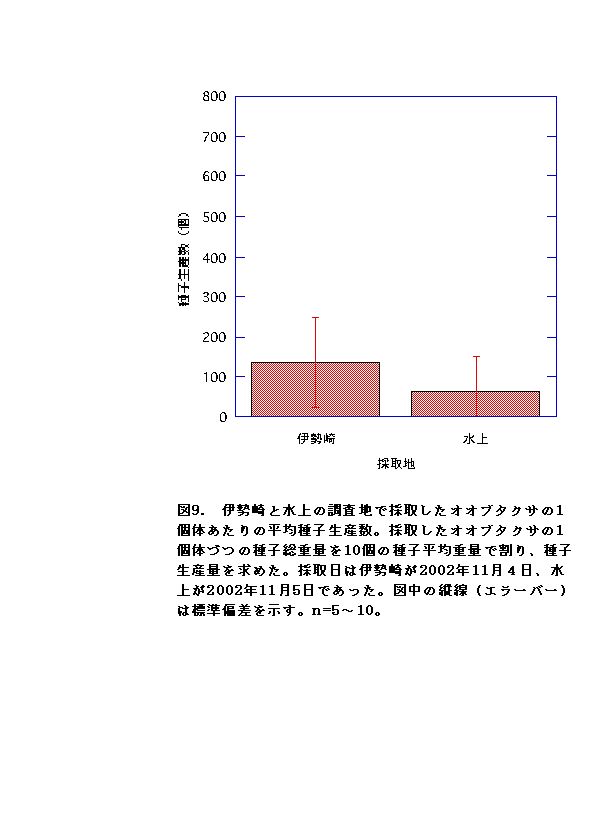
種子生産については、伊勢崎と水上の調査地点では、伊勢崎の方が水上より種子生産が多かった(図9)。サンプリング数が伊勢崎は5個体、水上は10個体と少なかったため、さらに検証が必要である。
実験 異なる窒素態比での栽培下における伊勢崎・水上両地点に生育するオオブタクサの生長・種子生産比較
2回の実験を通じて、オオブタクサの生長に対するアンモニア態窒素と硝酸態窒素の効果には、地域個体群間変異があることが示唆された。
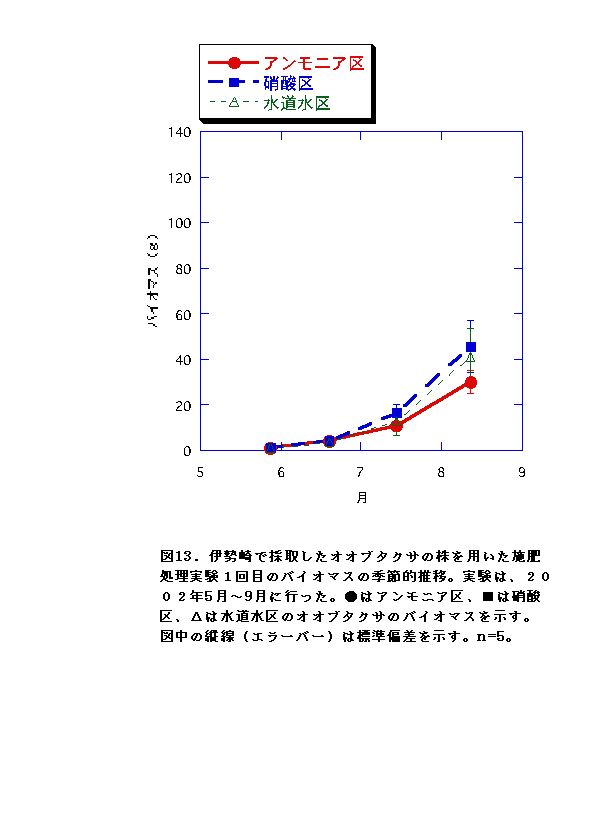
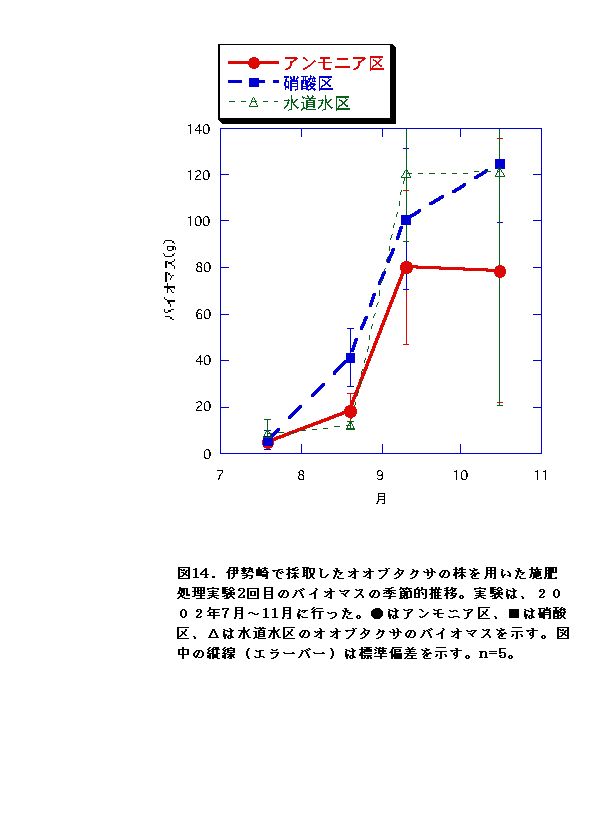
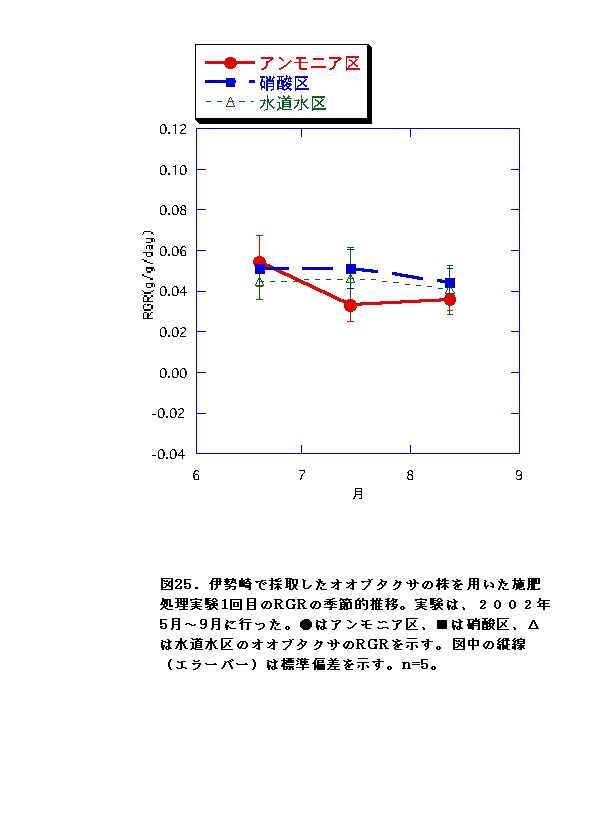
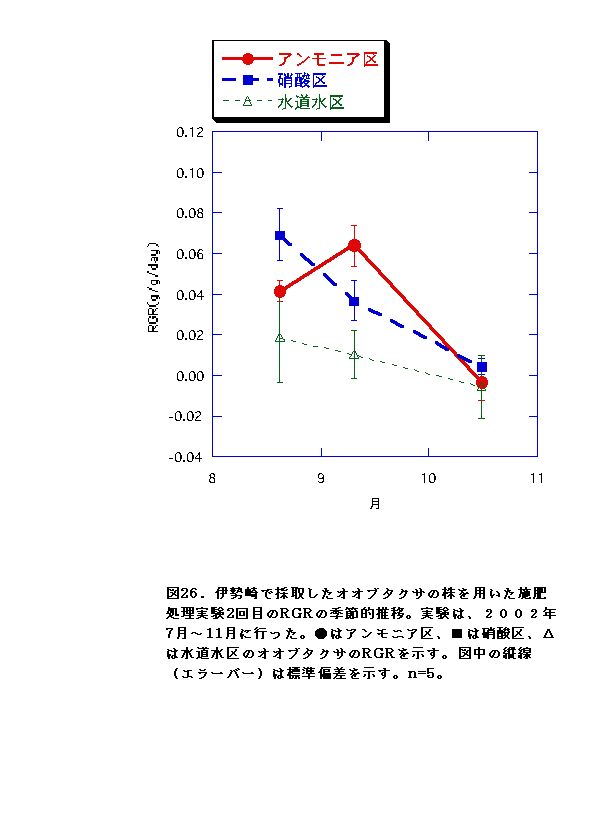
伊勢崎のオオブタクサにおいては、硝酸区と水道水区では、バイオマス、RGR共に顕著な差はなかった。一方、アンモニア区では、他の条件よりバイオマス、RGRが低くなった(図13、14、25、26)。これらのことから、アンモニア態窒素は伊勢崎のオオブタクサの生長を促進せず、むしろ生長を抑制する可能性があると言える。
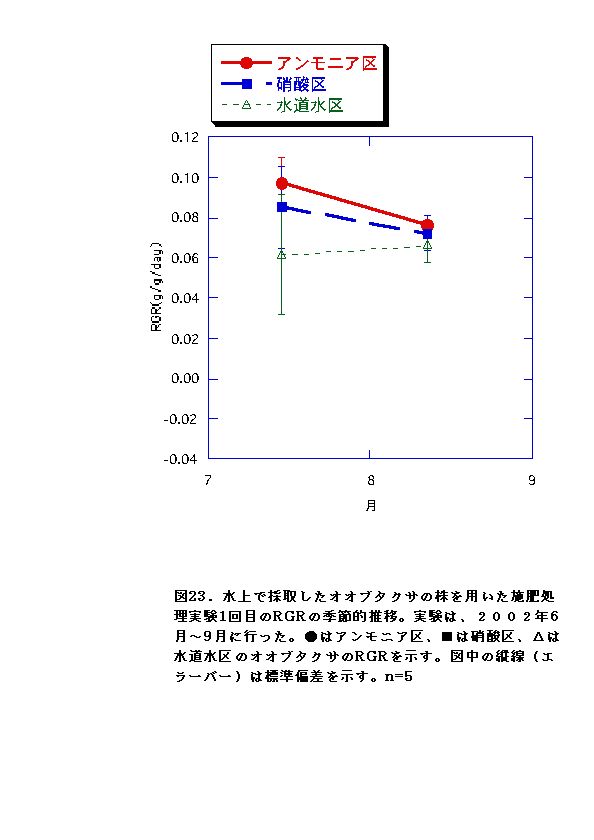
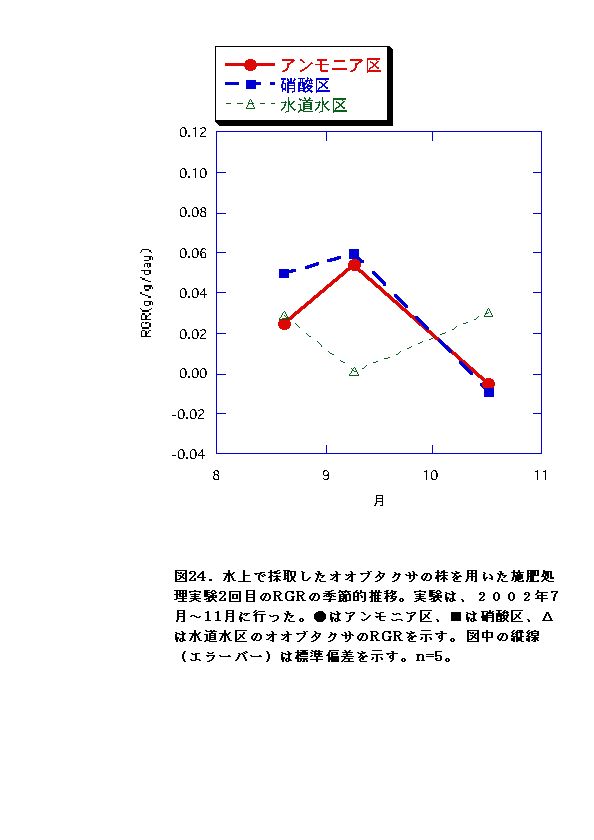
これに対して、水上のオオブタクサにおいては、処理区間でバイオマス,
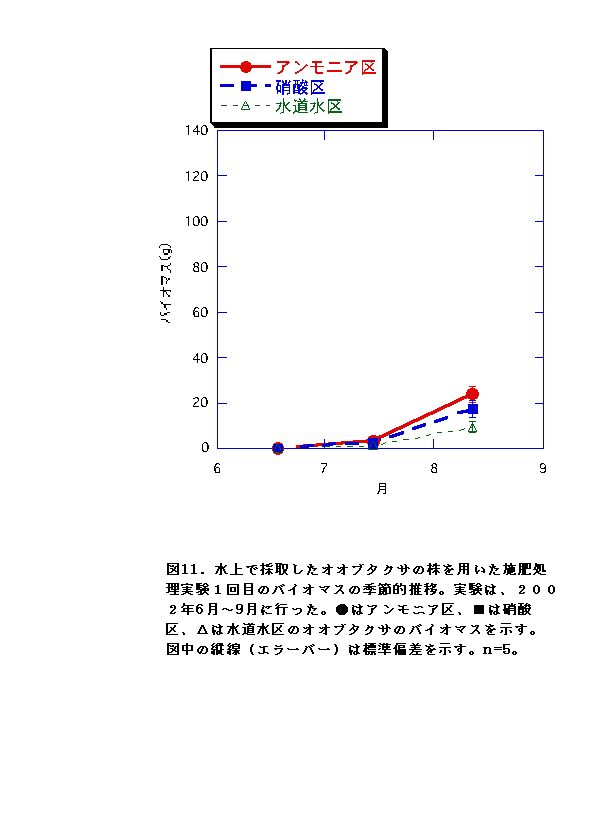
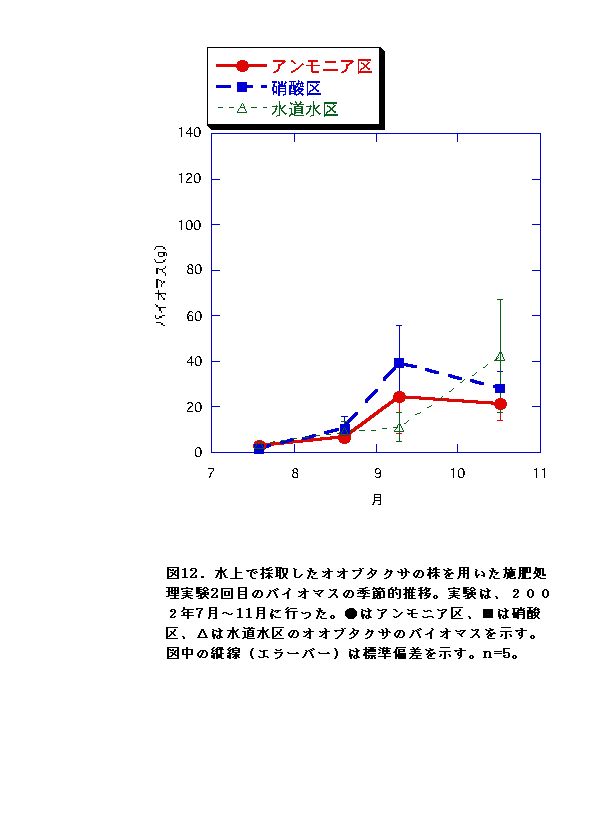
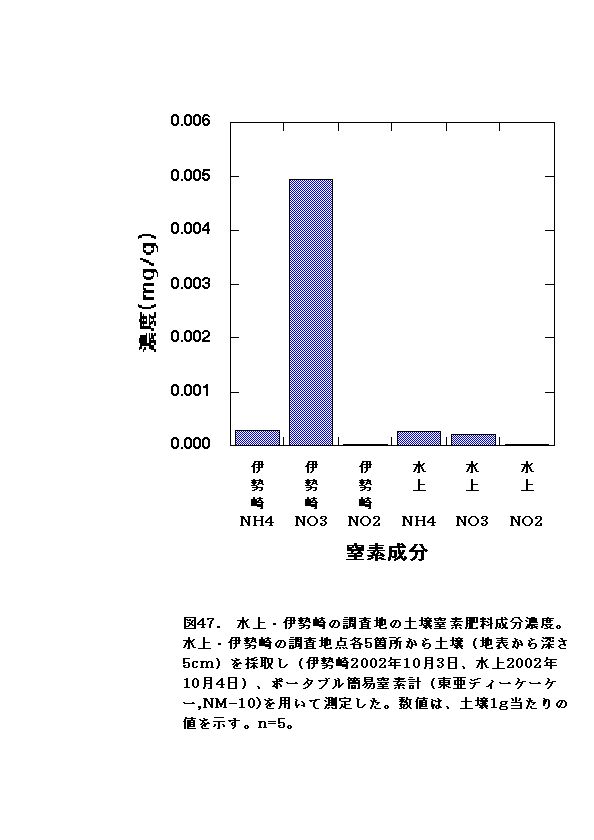
RGRに有意な差がないか、アンモニア区でむしろ大きくなる傾向が認められた(図11、12、23、24)。水上のオオブタクサ生育地では、土壌中の硝酸態窒素濃度が低く、相対的にはアンモニア態窒素比が高い状態となっていた(図47)。これらのことから、水上のオオブタクサにおいては、貧栄養・高アンモニア態窒素比に対する適応的進化が起こっている可能性があると言える。
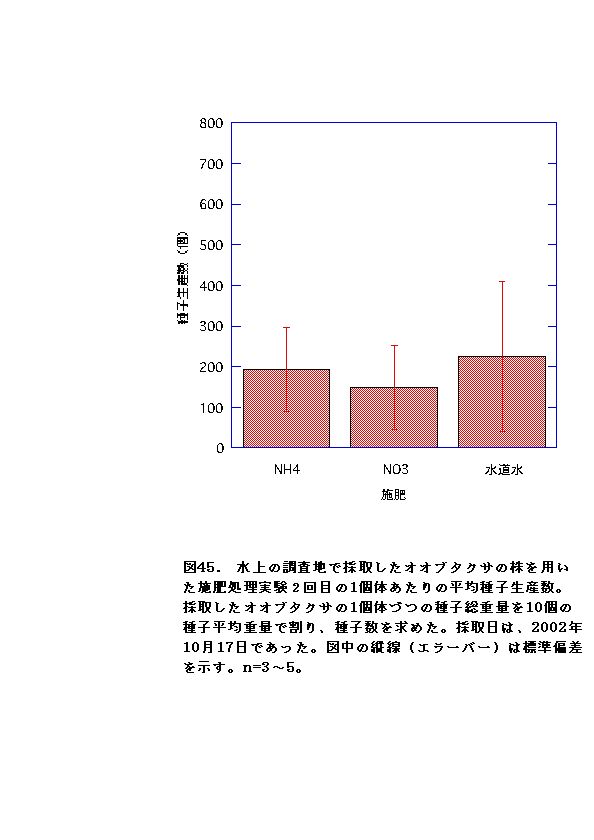
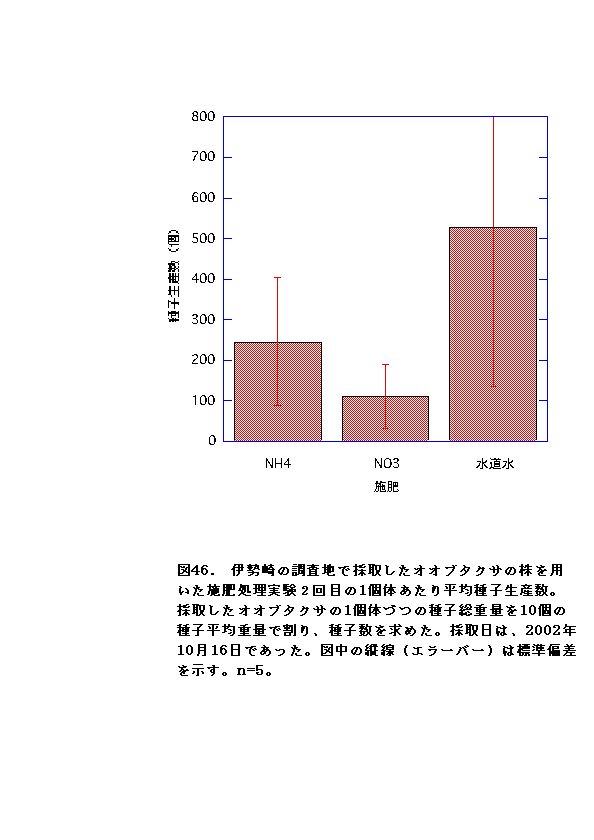
施肥実験における種子生産についてでは、水上から採取してきたオオブタクサでは、施肥によって種子生産に変化は無かった。一方伊勢崎から採取してきたオオブタクサでは、生長促進の効果はあったが、種子生産促進は効果が見られず、むしろ硝酸区では逆効果となった(図45、46・表7、8)。
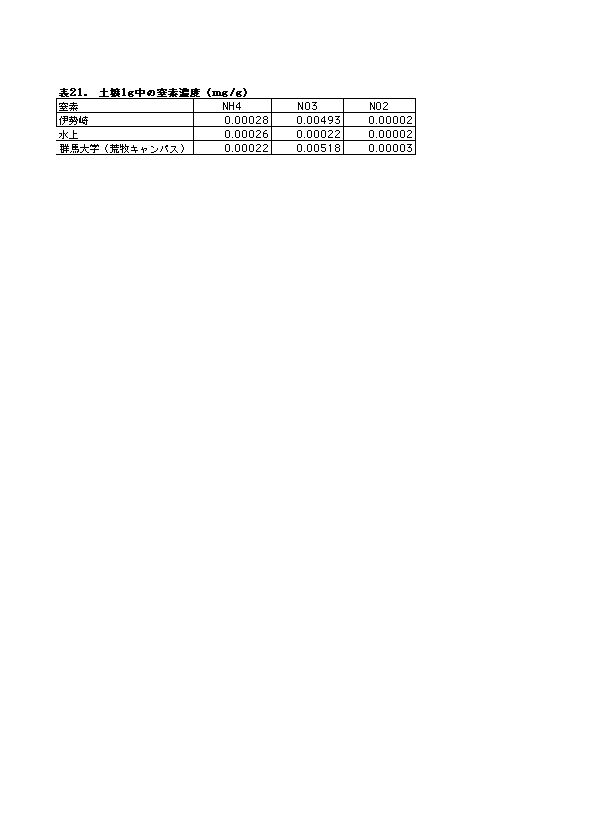
本研究では、実験開始時に水上に比べ、伊勢崎の初期の個体サイズが大きかったこと、栽培用に使った黒土にはすでに硝酸態窒素が多く含まれていた(0.005mg/g)こと(表21)、2回目の実験ではブタクサハムシに虫害を受けたことの3点が課題として残された。今後は虫害対策を強化するとともに、栽培用土壌窒素成分の見直し、伊勢崎のオオブタクサの株について実験初開始の時期を早くする等を考慮して、追試を行う必要がある。
実験¡ 水上・伊勢崎の気温とオオブタクサの生長の関係
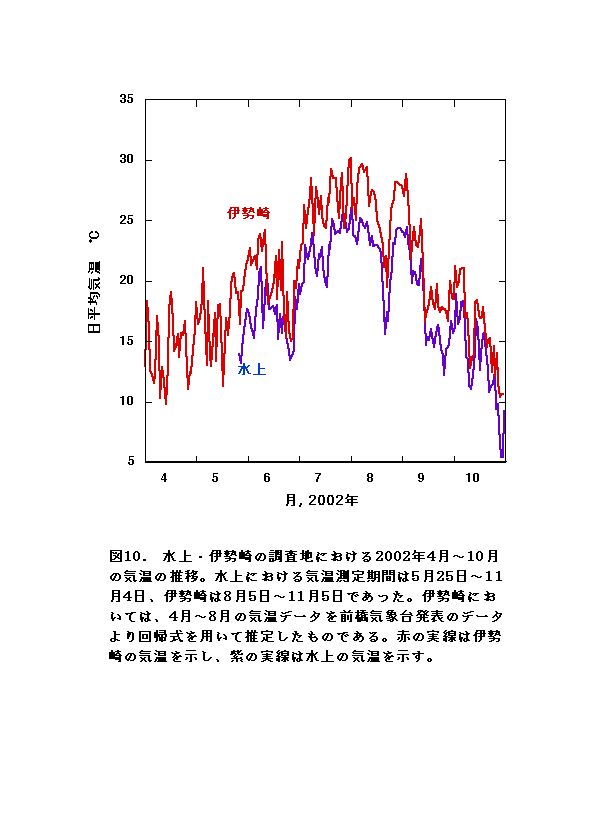
気温測定した5月から10月末までを通じ、日平均気温は伊勢崎の方が水上より2〜5℃高かった(図10)。気温の上昇が顕著に見られたのは、水上・伊勢崎両調査地点とも6月下旬から7月下旬の間だった。その期間、伊勢崎のオオブタクサではバイオマス増加が大きかったが、水上のオオブタクサでは顕著でなかった(図1)。このことから伊勢崎のオオブタクサは、夏期の気温の上昇と太陽光の増加を、効率的に生長に活かしていると考えられる。また、両調査地点とも気温が顕著に低下したのは、10月後半から11月上旬の間だった。その期間伊勢崎の調査地のオオブタクサは枯死していたものが多く見られたが、水上の調査地のオオブタクサは枯死しているものがほとんど見られなかった。このことから、水上の調査地のオオブタクサは、低温に適応している可能性があると考えられる。
実験¤ 水上・伊勢崎の土壌窒素肥料成分
両地の土壌の窒素成分で最も大きく異なったのは、硝酸態窒素濃度であった。伊勢崎の土壌は、水上の土壌の20倍以上も硝酸態窒素濃度が高かった(図47・表21)。このことから、伊勢崎のオオブタクサと水上のオオブタクサの生長・個体サイズの差が、両地の温度環境の違いだけではなく、土壌中の総窒素濃度および硝酸態窒素濃度の違いにも起因している可能性があると言える。
今後の課題としては、窒素測定をもっと多くの地点で行い、土壌窒素の比較実験を追試する必要がある。
実験¦前橋市内の主要幹線道路付近におけるハナダイコンの分布と光環境の関係
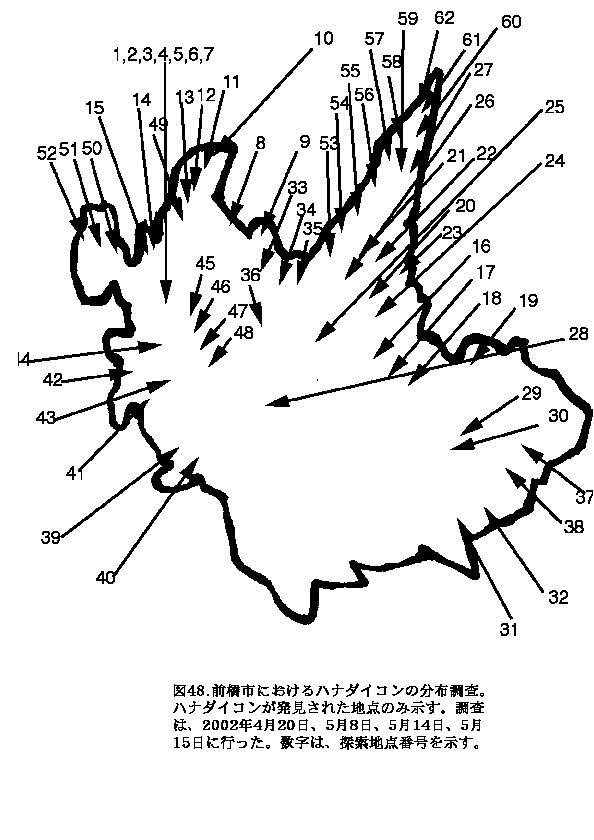
前橋市では、北部の方に分布が集中しており、南部にはほとんど分布が見られなかった(図48)。北部は、畑や、雑木林が多くあり、自生しやすい環境であったことが原因だと考えられる。南部は、市街地が多く、麦畑は若干見られたが、そこには自生していなかった。
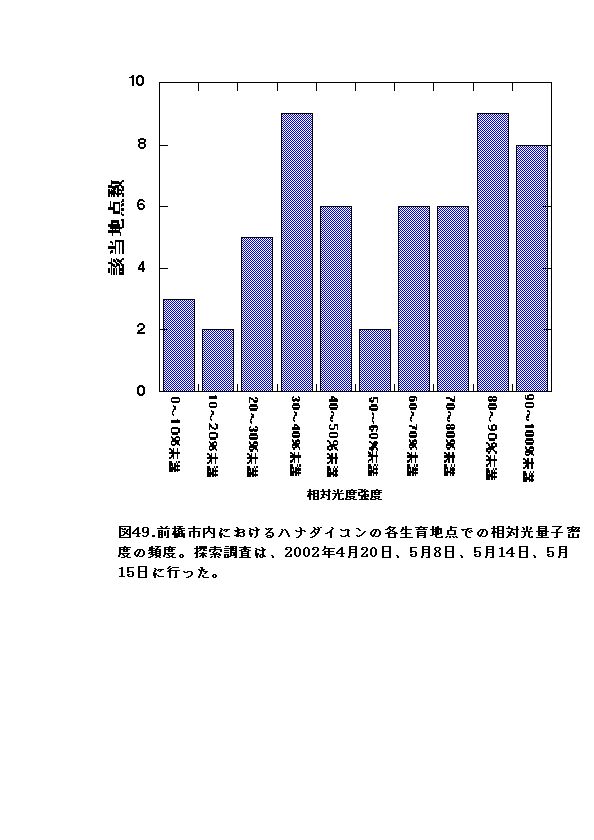
各分布地点の相対光量子密度は、大半が100〜30%の間にあり、10%以下のかなり暗い光度条件にはほとんどなかった(図49)。このことから、ハナダイコンは明るい所を好んで生育し、暗い所を嫌う特性があると考えられる。このことは、次節の実験によりさらに明確に検証がなされた。
実験© 人工被陰下におけるハナダイコンの栽培実験
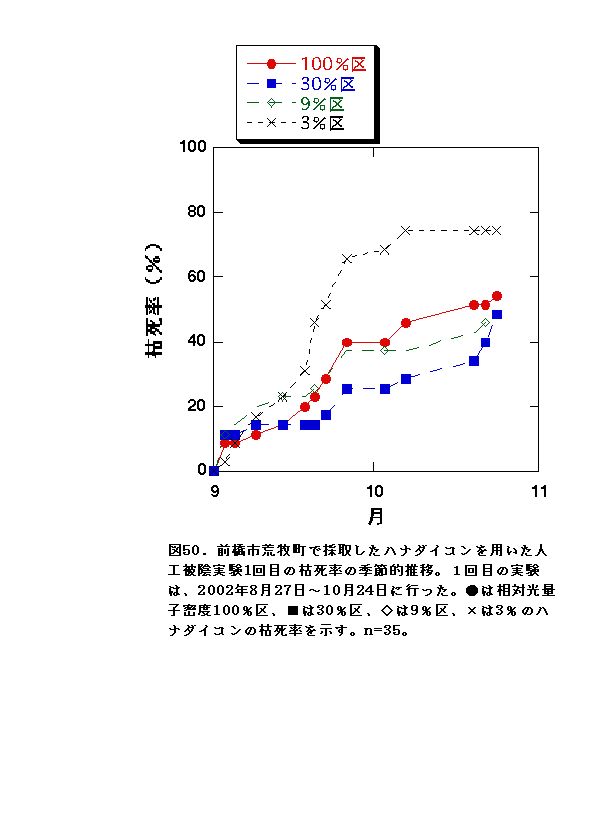
1回目の実験では、3%区での枯死率が最大となった(図50)。このことは、野外においてハナダイコンの分布地の相対光量子密度の大半が10%以上であったことと整合性がある。すなわち、ハナダイコンは10%以下の相対光量子密度では枯死率が増大するために、分布しにくいと考えられる。
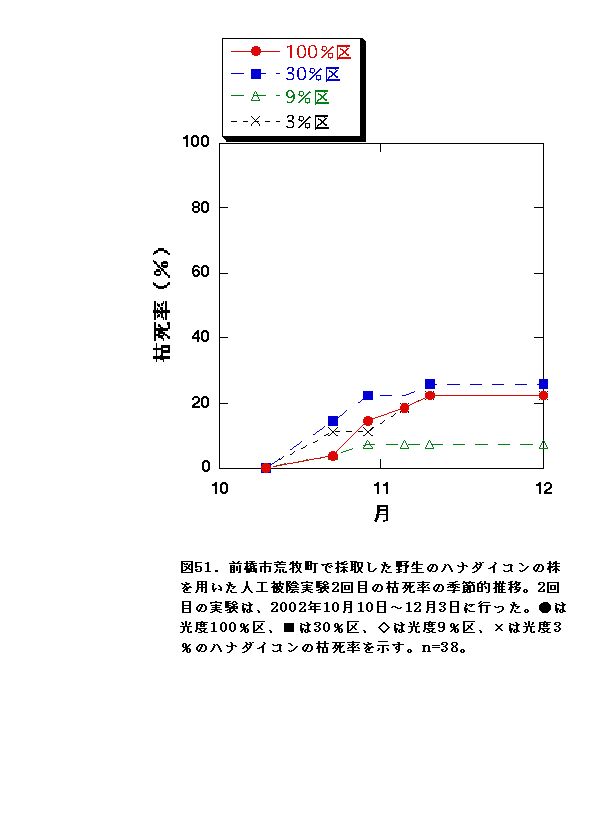
2回目の実験では、どの光度条件も、枯死率に顕著な差が無く、全体的に1回目の実験と比べて枯死したものが少なかった(図51)。この結果は、9月に開始した1回目の実験と10月に開始した2回目の実験の気温条件の差が要因だと考えられる。気温が高いと一般的に植物は呼吸の量が増え、低いと呼吸が減る性質がある。1回目の実験では、2回目の実験よりも高温のため植物の呼吸が多かったため枯れてしまい、2回目の実験では1回目の実験よりも低温だったため枯死する個体が少なくなったと考えられる。
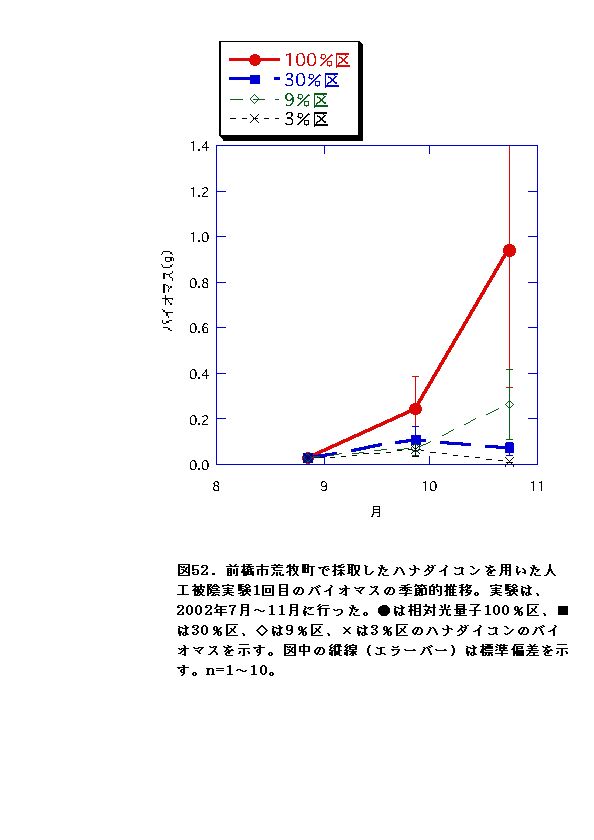
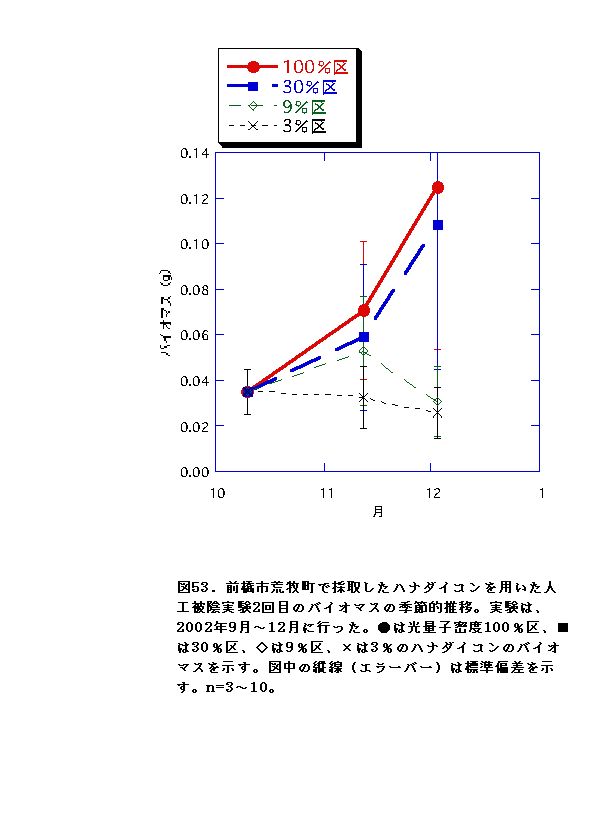
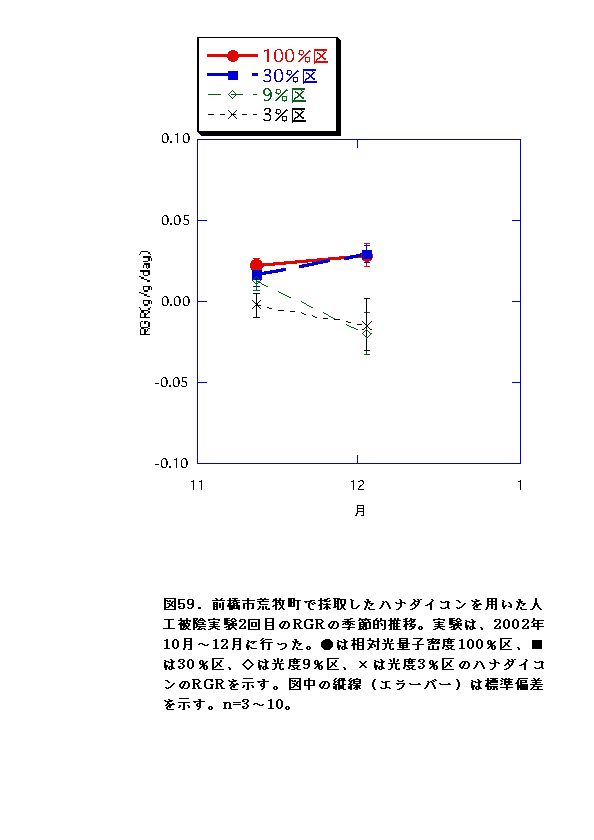
2回の実験を通じて、100%区でバイオマス・RGRがおおむね常に最大となり、3%区で最低となった(図52、53、58、59)。このことから、ハナダイコンは暗い環境下では生き残った個体であっても良好な生長は不可能であると言え、野外における本種の分布パターンの成因の一つであると考えられる。
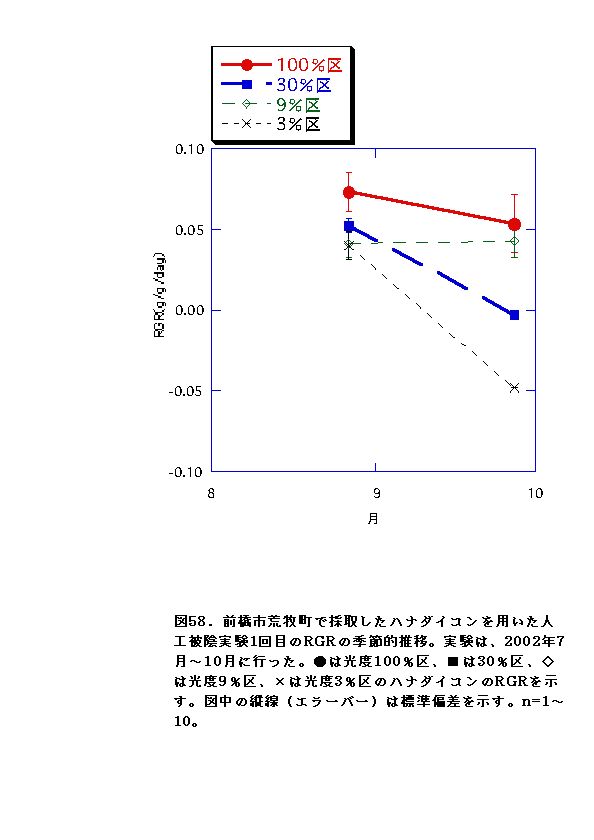
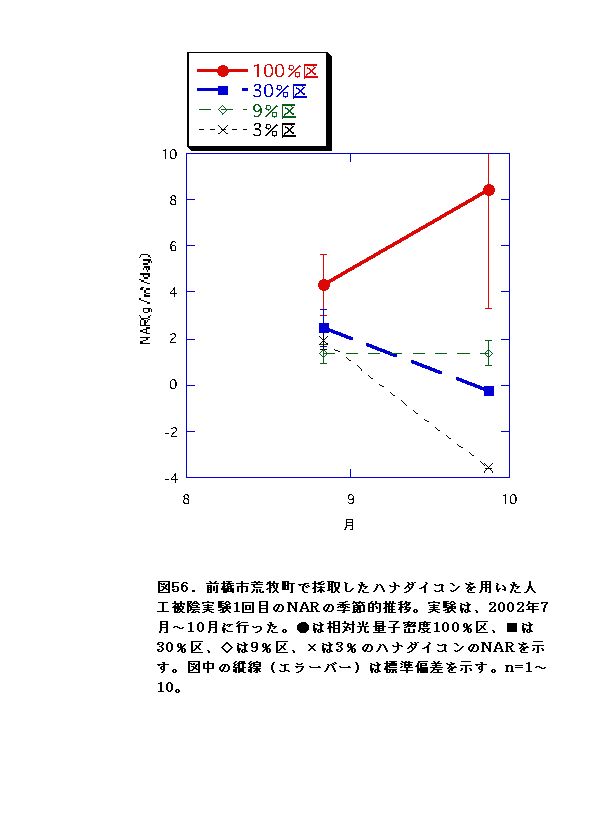
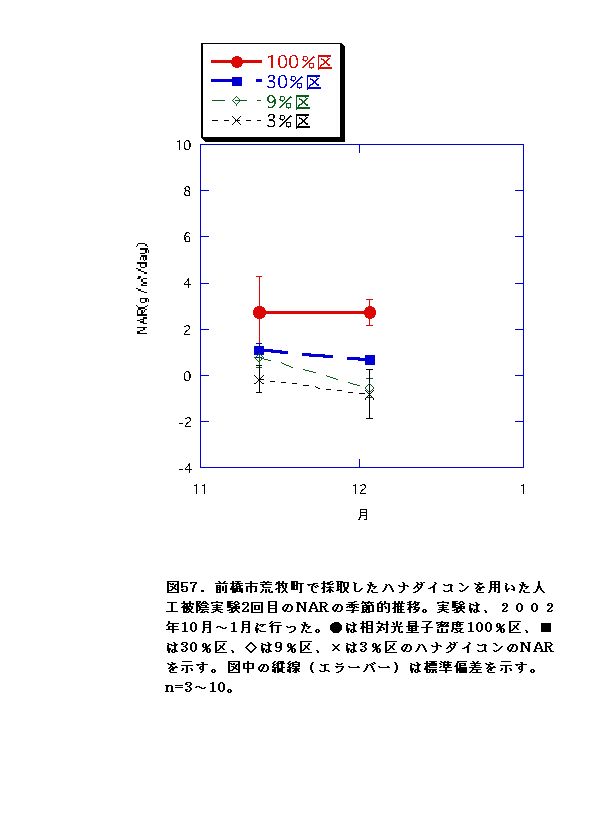
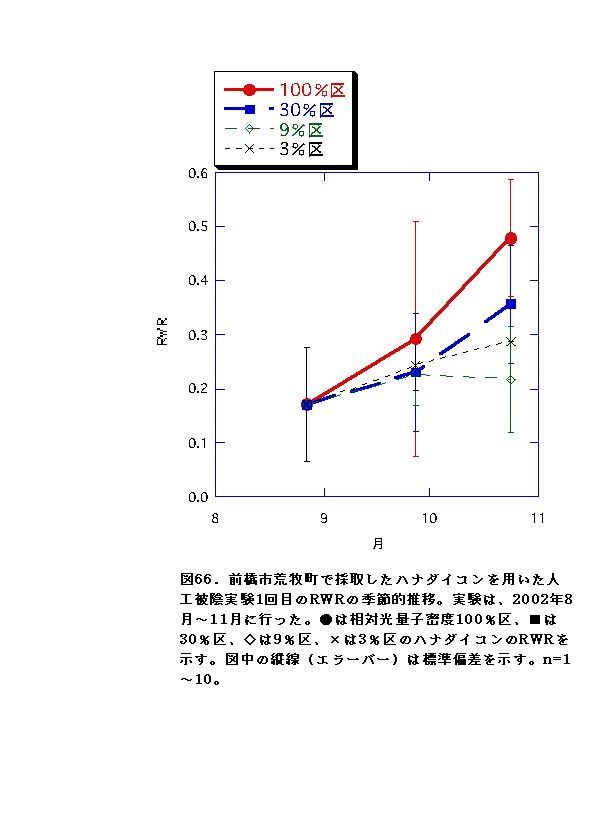
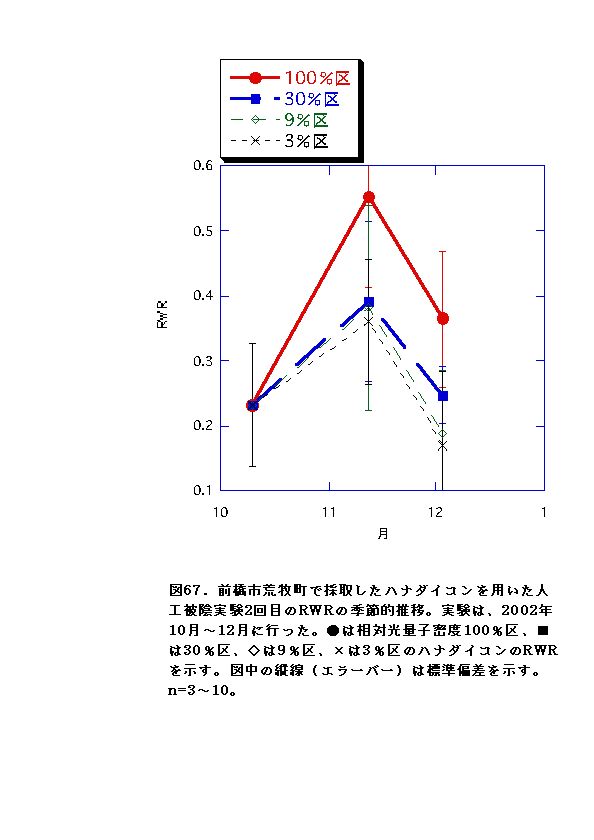
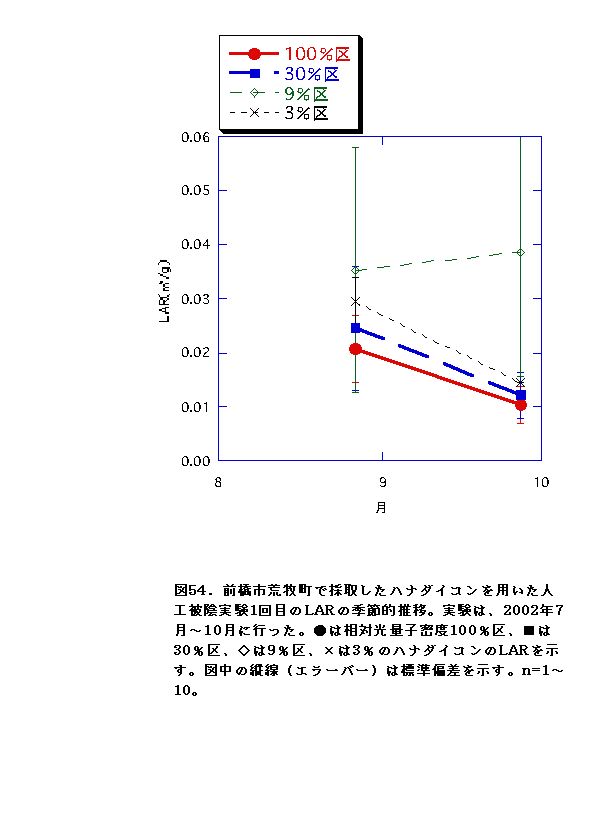
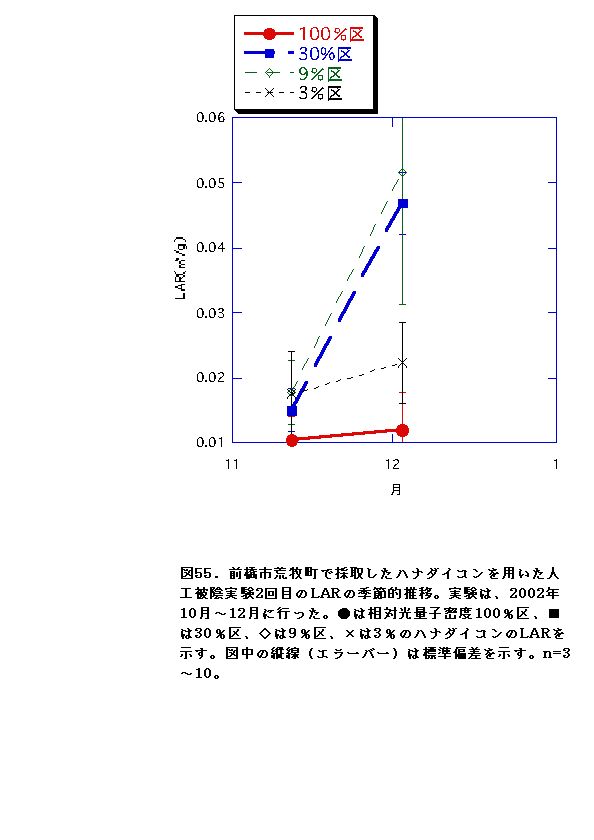
NARとRWRは、相対光量子密度が高いほど大きくなった(図56、57、66、67)。これは、光強度が大きいほど光合成が高くなると同時に葉の温度が上昇するため、より水分が必要になるので、根の生長が促進されるためと考えられる。またLARは9%区または30%区でで最大となり、3%区ではむしろ低下した(図54、55)。一般に植物は暗い生育環境下ほど葉面積を増やして光合成生産を高めようとするとされているが、限界はある。ハナダイコンにおいては、3%がその限界を超えていると考えられる。以上より、ハナダイコンが良好な生育・生存を行える限界光強度は、相対光量子密度で9%程度であり、その野外における分布パターンは、光強度により形成されるものであると考えられる。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}